深度解析医学证据,DeepEvidence为你支撑决策
小鼠双微体 2(MDM2) 的药理学靶向作用是血液恶性肿瘤中p53 野生型治疗性再激活最具前景服的策略之一。MDM2-p53自我调节环路是细胞应激反应的核心调控枢纽。在髓系肿瘤(包括急性髓系白血病和骨髓增殖性肿瘤)中,p53通常未被突变,但其功能受到MDM2过表达及致癌信号(尤其是JAK-STAT通路激活)的抑制。过去十年中,多代MDM2抑制剂相继问世,将结构和机制上的突破转化为临床研究,并在给药方案、血液学毒性、生物标志物驱动的患者选择及耐药机制(包括TP53突变克隆的选择性扩增)方面积累了关键经验。尽管在AML中的III期临床试验结果为阴性,但近期在骨髓纤维化中的研究显示,MDM2抑制剂能带来有临床意义的脾脏、症状和分子学反应,支持其在TP53野生型背景下的疾病修饰潜力。适应性平台设计以及联合JAK抑制剂、BCL-2拮抗剂、干扰素等合理组合策略进一步优化了治疗方案。新兴的MDM2降解剂和下一代药物旨在克服反馈性限制并改善治疗指数。
《Leukemia》近日发表综述,整合了机制基础、临床开发、耐药生物学及未来方向,强调了数十年的基础科学研究如何将p53再激活重塑为髓系疾病中的一种精准治疗策略。

概述
鼠双微体 2(MDM2) 的药理学靶向作用已成为现代血液肿瘤学概念上最重要和具有临床指导意义的进展之一。该领域扎根于MDM2-TP53自我调节环路作为细胞应激反应主调控因子的基础发现,现已从结构和生化研究进展为激活内源性抑癌功能的临床可行策略。在髓系恶性肿瘤(尤其是MPN和AML)中,TP53通常保持野生型但功能受抑,MDM2抑制为治疗性利用这一完整但受抑制的p53通路提供了独特机会。过去十年中,多代MDM2抑制剂将这些原理转化为临床实践,在靶点生物学、给药方案、毒性管理、克隆进化及生物标志物驱动的患者选择方面积累了关键经验。同时,新兴数据揭示MDM2是致癌信号、炎症信号和细胞因子驱动通路(尤其是MPN中的JAK-STAT信号)的汇聚点,从而使MDM2抑制不仅是一种细胞毒性策略,更可能成为一种疾病修饰性干预手段。
在本综述中,作者整合了MDM2-p53轴的机制基础与MDM2抑制剂的发展历程,总结了在髓系肿瘤中的关键临床经验,并讨论了耐药机制、安全性考量及未来方向,包括合理的联合策略和下一代方法(如MDM2降解剂)。这些见解共同描绘了数十年的基础生物学研究如何最终形成一个治疗范式,并持续重塑我们对髓系疾病中p53再激活的理解。
机制基础:细胞稳态中的MDM2-TP53轴
MDM2的结构与功能架构
MDM2是抑癌基因p53的主要负调控因子,也是一个整合结构特异性、翻译后修饰和亚细胞运输以控制细胞命运决定的中枢信号枢纽。MDM2癌蛋白由多个功能特化的结构域组成,每个结构域都为其调控 versatility 做出贡献。其N末端结构域包含p53结合界面,形成一个深疏水裂隙,用于容纳p53的双亲性α螺旋。高分辨率晶体学研究表明,p53的三个保守残基(Phe19、Trp23和Leu26)插入此口袋,直接遮蔽p53的反式激活域,从而抑制下游靶基因的转录激活。
MDM2中央部分包含一个酸性结构域,随后是一个锌指基序,介导与p53之外的其他蛋白相互作用,尤其是与核糖体蛋白L5和L11。这些相互作用将MDM2活性与核糖体生物合成及核仁应激感应相偶联。影响锌配位半胱氨酸残基(如C305F、C308Y)的癌症相关突变会破坏核糖体蛋白结合,改变MDM2亚细胞定位,并损害有效的p53降解,突显该结构域在维持调控保真度中的重要性。C末端的RING指结构域赋予E3泛素连接酶活性,使MDM2能够招募E2泛素结合酶,催化p53 C末端赖氨酸残基的单泛素化和多泛素化,从而将p53靶向蛋白酶体降解。此外,MDM2具有内在的核定位和核输出信号,使其能够作为核质穿梭蛋白,将p53从细胞核运至细胞质蛋白酶体,为p53抑制增加了空间层面的调控(图1)。
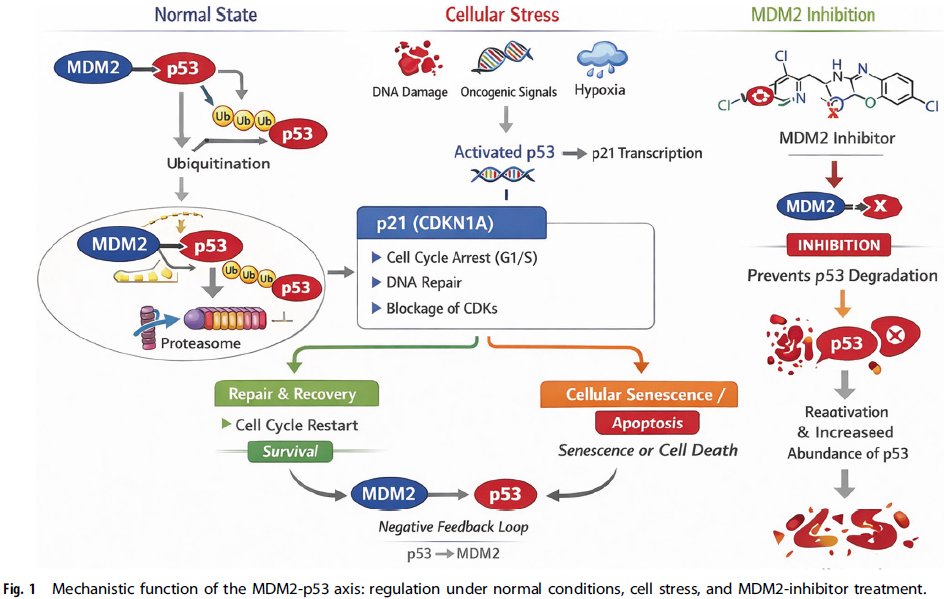
自我调节反馈环路
MDM2-p53相互作用嵌入在哺乳动物细胞中调控最严密的自我调节反馈环路之一。在基础条件下,通过MDM2介导的持续泛素化和降解,p53维持在较低稳态水平。当细胞遭遇应激(如DNA损伤、癌基因激活、缺氧或炎症信号)时,p53被稳定并具有转录活性,诱导广泛的应激反应程序。其最早和最稳健的转录靶点之一正是MDM2本身,从而形成了一个负反馈环路:p53驱动其自身抑制剂的表达。新合成的MDM2随后通过多个部分冗余的机制减弱p53活性:直接结合遮蔽反式激活域、增强核输出、以及泛素依赖性蛋白酶体降解(图1)。
一旦激发应激得到解决,该反馈环路可确保p53信号快速终止,从而防止不适当或持续的p53活性,后者可能引发过度的细胞周期阻滞或凋亡。通过翻译后修饰(磷酸化、乙酰化或neddylation)破坏这一平衡,可暂时解偶联该复合物,导致p53稳定并选择性转录下游效应基因。其中,CDKN1A(p21)作为关键的分子变阻器,调控可逆的细胞周期阻滞、衰老和凋亡承诺之间的平衡。持续的p21诱导有利于持久的生长停滞或衰老,而不足或短暂的p21激活则允许由BAX、PUMA和NOXA介导的p53依赖性凋亡程序的启动。
MDM2 的非p53依赖性功能
除了其在p53调控中的经典作用外,MDM2还发挥多种p53非依赖性致癌功能,极大地拓宽了其生物学和临床价值。MDM2与细胞周期进展和存活的关键调控因子(包括RB、E2F-1和FOXO3A)相互作用,从而即使在p53缺陷的环境中也促进增殖和凋亡抵抗。此外,MDM2通过稳定VEGF mRNA和在缺氧条件下与HIF-1α功能性合作,促进肿瘤血管生成,将代谢应激反应与新血管生成联系起来。
新兴证据进一步表明MDM2参与炎症和细胞因子驱动的恶性肿瘤。在MPN中,JAK2致癌信号下游的JAK-STAT通路组成性激活已被证明可在转录和翻译后水平上调MDM2的表达和活性。这加强了造血祖细胞中的p53抑制,并在功能上将炎症信号与受损的抑癌控制相偶联。因此,MDM2充当了致癌、炎症和应激反应通路的汇聚点,为在MPN及其他以完整但功能受抑的p53为特征的恶性肿瘤中,单独或联合JAK-STAT抑制剂靶向MDM2的治疗策略提供了机制依据。
MDM2抑制剂的发展历程
第一代抑制剂:Nutlin时代
现代药理学MDM2抑制的时代始于nutlin类小分子的发现,它首次令人信服地证明,像MDM2-p53这样具有挑战性的蛋白-蛋白相互作用可以被治疗性靶向。Nutlins是顺式咪唑啉衍生物,通过合理设计占据MDM2上通常容纳p53反式激活域的疏水口袋,从而模拟p53的关键残基Phe19、Trp23和Leu26,阻止p53结合。早期先导化合物如Nutlin-3a源于结构引导的优化,并成为广泛使用的化学探针,用于验证选择性破坏MDM2-p53相互作用可导致野生型p53快速稳定、典型p53靶基因的转录诱导、细胞周期阻滞及癌细胞凋亡。
然而第一代nutlins存在药代动力学和代谢缺陷,阻碍了直接临床转化,促使人们进行广泛的药物化学研究以提高效力、口服生物利用度和体内暴露量。这些优化工作最终促成了RG7112的发现,它是一种第二代nutlin衍生物,也是首个进入临床评估的MDM2抑制剂。RG7112保留了核心的咪唑啉骨架,但进行了关键的结构改进,包括4,5-二甲基取代,增强了代谢稳定性和结合亲和力,同时保留了经典的nutlin结合模式。临床前研究中,RG7112显示出对MDM2的纳摩尔级亲和力、p53信号的 robust 激活以及在异种移植模型中的相关抗肿瘤活性,尤其是在MDM2过表达且TP53完整的肿瘤中。RG7112进入早期临床试验,为药理学再激活p53在患者中的可行性提供了关键的概念验证。在复发/难治性白血病中,RG7112诱导了p53稳定、p53靶基因的转录上调以及可测量的抗白血病活性,包括在部分AML患者中观察到客观反应。同时,这些研究揭示了一个狭窄的治疗窗口。剂量限制性毒性主要是靶向性的造血抑制,尤其是血小板减少和中性粒细胞减少,反映了MDM2在正常造血过程中抑制p53的生理作用。胃肠道毒性和较大的药片负担进一步使剂量递增复杂化。尽管如此,nutlin时代对该领域具有变革性意义。RG7112最终验证了MDM2是人类癌症中可成药的靶点,并将p53再激活确立为临床相关的治疗策略。重要的是,第一代nutlins的毒性特征和药理学缺陷提供了宝贵的机制和转化见解,直接指导了具有改进效力、选择性和给药特征的下一代MDM2抑制剂的设计,最终塑造了后续MDM2靶向药物开发的浪潮。
第二代进展:Idasanutlin
Idasanutlin (RG7388; RO5503781)是作为第二代MDM2抑制剂开发的,其明确目标是克服第一代nutlin化合物(如RG7112)的药理学和耐受性限制。RG7112确立了p53再激活的临床概念验证,但其需要持续的高全身暴露量,导致剂量限制性血液学毒性,限制了治疗灵活性。Idasanutlin源于一项集中的药物化学研究,通过战略性骨架重新设计,用结构不同的吡咯烷基化学型取代了顺式咪唑啉nutlin核心,同时保持了对MDM2 p53相互作用口袋的高亲和力结合,从而提高了效力、选择性和药代动力学行为。
这种骨架优化产生了一种化合物,其生化效力和细胞活性相对于RG7112显著增强,能够在较低浓度下实现野生型p53的稳健稳定化和典型p53靶基因的转录诱导。重要的是,idasanutlin表现出改善的口服生物利用度和更有利的药代动力学特征,转化为在预测临床更可耐受的暴露水平下的有效体内靶点结合。这些特性共同将idasanutlin定义为一种药理学上更易处理的MDM2拮抗剂,适合在短期概念验证给药之外进行探索。
与idasanutlin相关的一个关键概念进展在于认识到持久的抗肿瘤疗效不需要连续的MDM2抑制。详细的临床前药代动力学-药效动力学建模揭示了药物暴露、p53稳定和凋亡执行之间的时间脱节。体外脉冲暴露实验表明,短暂的idasanutlin治疗足以不可逆地使肿瘤细胞走向凋亡,最大细胞死亡发生在药物洗脱之后。相应地,体内异种移植研究表明,间歇性给药方案(如每周一次或短期给药)产生的肿瘤生长抑制作用与连续每日给药相当,尽管累积药物暴露量减少,这些发现具有直接的转化意义。预计间歇性给药可在保留抗肿瘤疗效的同时,减轻对正常组织(尤其是造血区室)的靶向毒性,因为在这些组织中,MDM2介导的生理性p53抑制对于祖细胞存活至关重要。因此,idasanutlin的早期临床开发优先考虑了28天周期内的间歇性方案,这不同于RG7112采用的近乎连续暴露模式。这种方案设计代表了第二代MDM2抑制剂发展的一个决定性特征,整合了机制见解与实用的临床试验设计。
在临床上,idasanutlin在实体瘤和血液系统恶性肿瘤(包括AML)中进行了多项1期研究,在TP53野生型疾病中观察到了可预测的p53通路激活。这些研究进一步细化了idasanutlin的安全性特征,确认血液学抑制和胃肠道效应仍是主要的靶向毒性,但通常可通过间歇性给药策略进行管理。重要的是,一项整合了三个1期试验数据的汇总浓度-QTc分析显示,无暴露依赖性QT延长,有效消除了慢性或联合给药的一个关键潜在风险。Idasanutlin改善的治疗指数使得能够系统探索合理的联合策略,尤其是在AML中,假设MDM2抑制可与增强凋亡启动或抑制代偿性生存通路的药物协同作用。尽管后续的后期临床结果好坏参半,但idasanutlin仍然代表了MDM2抑制剂发展过程中的一个关键成熟点。它证明,结构引导的药物化学结合机制指导的给药策略,可以显著提高p53再激活作为一种治疗方法的临床可行性。在此过程中,idasanutlin确立了继续指导更新型MDM2拮抗剂和p53通路靶向联合方案在白血病及其他领域发展的关键原则。
第三代高效抑制剂:Siremadlin和Navtamadlin
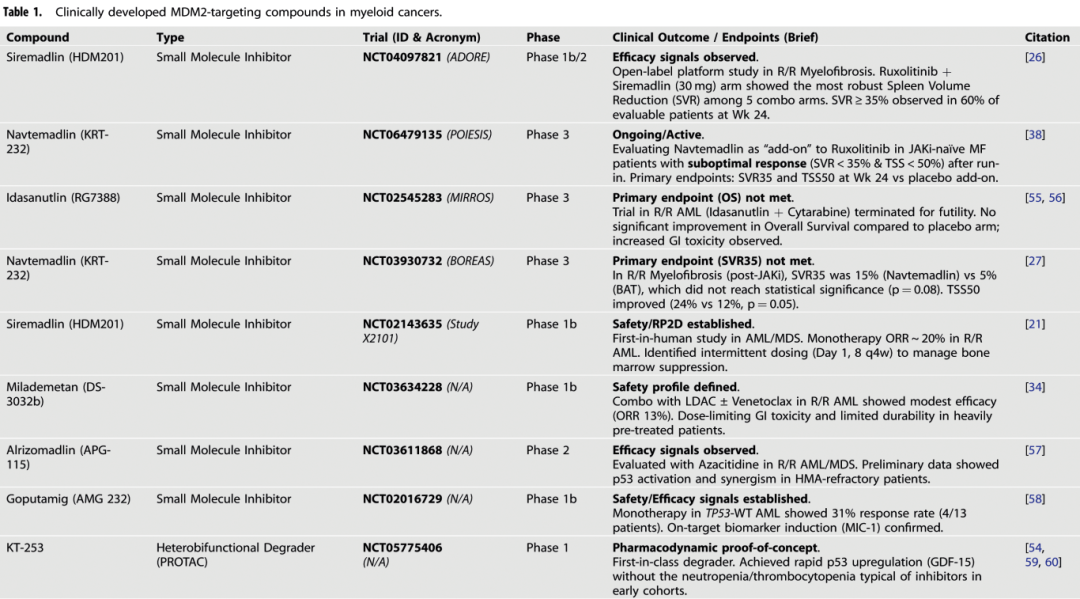
当前一代的MDM2抑制剂是结构生物学、药物化学和临床经验长期研究的结晶。Siremadlin (HDM201, NVP-HDM201)对MDM2具有皮摩尔级亲和力,对MDM4的选择性超过10,000倍。这种选择性特征使得治疗方法更具靶向性,脱靶效应减少。
Siremadlin的临床开发主要集中在实体瘤,包括脂肪肉瘤和骨髓纤维化等多种恶性肿瘤的II期研究。值得注意的是,siremadlin在AML中的开发因血液学毒性挑战而中止。然而最近的研究显示其在TP53野生型的慢性淋巴细胞白血病中具有令人鼓舞的活性。
Navtamadlin (KRT-232, APG-115)是另一种具有独特药理学特性的同类最佳MDM2抑制剂。该化合物在MF中显示出显著的临床活性,促使其进入III期BOREAS试验进行评估。Navtamadlin在MF中的成功是该领域的一个分水岭,证明了MDM2 抑制剂 可以在血液系统恶性肿瘤中实现有临床意义的结局。
髓系肿瘤中的开创性MDM2研究
MPN生物学中的基础发现
Ronald Hoffman的研究工作在将p53信号失调定义为MPN中一个核心且可治疗性利用的弱点方面起到了关键作用。通过整合的机制和转化研究,建立了一个框架,其中疾病起始的造血干细胞和祖细胞(而非成熟的效应细胞群)被确定为关键的治疗靶点。在MPN患者的CD34+细胞中发现了MDM2过表达,与真性红细胞增多症和正常对照相比,原发性MF中的水平最高。值得注意的是,这发生在TP53基本完整的背景下,表明是功能性抑制而非遗传性失活。这些发现将MDM2定位为疾病进展的生物标志物和慢性期MPN中的合理治疗靶点。药理学拮抗MDM2被证明可选择性地恢复恶性祖细胞中的p53活性。Nutlin类化合物优先抑制源自PV CD34+细胞的红系和髓系集落形成,同时基本不影响正常造血。单个集落的基因分型显示,JAK2V617F阳性祖细胞(尤其是杂合克隆)被选择性清除,这与突变细胞对MDM2介导的p53抑制的依赖性增加一致。这些效应与p53稳定及下游细胞周期和凋亡调控因子(包括p21、PUMA和BAX)的诱导相关。联合治疗的机制基础源于将MDM2拮抗剂与聚乙二醇化干扰素-α2a联合的研究。观察到p53通路的互补激活:干扰素通过STAT1和p38 MAPK信号增强p53转录,而MDM2抑制阻止p53降解。在亚治疗剂量下,这种组合协同增强了MPN CD34+细胞的凋亡,并减少了JAK2V617F阳性集落形成,支持了一种克隆耗竭而非单纯细胞减少的策略。这些临床前发现已转化为口服MDM2抑制剂的临床研究。在一项研究者发起的idasanutlin治疗高危、治疗难治性PV和原发性血小板增多症的1期研究中,观察到了靶向性p53激活、血液学和症状反应以及JAK2V617F等位基因负荷的降低,并在添加低剂量干扰素-α2a后活性增强。生物标志物驱动的转化策略是这项工作的核心。药效动力学指标,包括p53靶基因诱导、JAK2V617F等位基因 burden 的连续评估以及骨髓组织病理学,使得能够评估超越外周血计数正常化的生物学反应。同时,开发了患者来源的异种移植模型来研究对分层组织的MPN干细胞群体的治疗效应,为识别持久疾病修饰所需的细胞区室提供了关键的临床前平台。
总之,这些研究确立了功能性p53抑制作为慢性期MPN的一个定义特征,并为旨在实现持续克隆和组织学反应的干细胞定向治疗策略提供了转化蓝图。
Hoffman及其同事率先开展的联合研究,为MPN中合理的疾病修饰策略奠定了关键的临床前基础。使用来自PV和原发性MF患者的原代CD34+细胞,他们证明口服生物可利用的MDM2拮抗剂RG7112,特别是与聚乙二醇化干扰素-α2a联合使用时,对恶性造血祖细胞和干细胞区室发挥出强效且选择性的作用。在能够保护正常造血的亚优浓度下,这种组合显著减少了CFU-GM和BFU-E集落形成,并优先耗竭了JAK2V617F阳性祖细胞,杂合突变集落的减少尤为显著。机制上,这些效应与p53-MDM2相互作用的破坏有关,导致p53信号恢复并选择性地在MPN CD34+细胞中诱导凋亡。重要的是,功能性干细胞分析显示,短期体外暴露于RG7112和聚乙二醇化干扰素-α2a会显著削弱MPN CD34+细胞在免疫缺陷NSG小鼠中植入和 repopulate 的能力,同时伴随着体内JAK2V617F等位基因 burden 的大幅降低。这些数据提供了直接证据,表明联合 再激活p53通路和干扰素信号可以靶向MPN中的疾病起始细胞,从而为旨在克隆耗竭而非单纯细胞减少的低剂量联合方法提供了强有力的生物学依据——这一见解随后指导了基于MDM2抑制剂的联合方案进入早期临床研究。
当代临床进展
MPN中的第三代MDM2靶向治疗正越来越多地从概念验证性抑制剂转向更有效、方案更灵活的药物,旨在加深脾脏和症状反应,并可能改变疾病生物学。Siremadlin正在适应性的ADORE平台内的芦可替尼联合策略中进行探索,早期信号表明,将p53再激活与JAK抑制相结合可以增强MF中的临床活性。与此同时,Navtamadlin (KRT-232)已进入MF的后期开发阶段,包括在JAK抑制剂复发/难治性疾病中的单药注册研究(BOREAS)以及全球III期POIESIS试验,后者专门评估navtamadlin作为芦可替尼的添加疗法,用于对芦可替尼反应欠佳的JAK抑制剂初治患者——这一设计与现实世界的临床决策相一致。展望未来,下一代MDM2抑制剂和新兴的MDM2靶向蛋白降解靶向嵌合体旨在通过从可逆阻断转向靶向MDM2消除来改善治疗指数,可能提供更持久的p53通路再激活,并随着这些模式进入早期临床转化而提供新的联合选择。
ADORE:临床试验设计的平台创新助力siremadlin联合治疗成功
ADORE(Assessment of Dual Combinations in Ruxolitinib Nonoptimal Responders with Myelofibrosis)研究是罕见血液癌症临床试验设计的重要进步。通过实施一项1b/2期开放标签、适应性平台架构,ADORE解决了在对芦可替尼反应欠佳的MF患者群体中测试多种研究性疗法的核心挑战(表1)。ADORE没有在各自需要独立入组、基础设施和对照臂的传统早期试验中依次评估候选药物,而是将芦可替尼作为治疗基石,并允许同时、协调地评估五种机制不同的药物:siremadlin、rineterkib、sabatolimab、crizanlizumab和NIS793。这种整合设计提高了临床开发的效率和科学产出,能够快速优先排序有前景的组合,同时最大限度地减轻有限患者群体的负担。
在第1部分入组的44名患者中,最大的队列接受了芦可替尼和MDM2抑制剂siremadlin的联合治疗(n=23)。这些患者的中位年龄为68岁,主要为原发性MF(65.2%),之前接受过大量的芦可替尼治疗(中位67.9周;在30mg siremadlin亚组中为165.1周)。基线时的分子分型显示,14名患者存在JAK2V617F突变,5名存在CALR突变,10名存在ASXL1、SRSF2、EZH2、IDH1/2或U2AF1的高危突变,反映了晚期MF特有的突变异质性。Siremadlin是一种有效的MDM2抑制剂,旨在 reactivate p53介导的凋亡,从而促进克隆消退。临床前数据表明,激活p53通路与JAK抑制相结合可能会将疾病生物学转向更深层次且可能具有疾病修饰作用的反应,这为该组合提供了机制依据。
芦可替尼联合siremadlin的安全性特征与MDM2抑制的预期效应以及MF人群潜在的造血脆弱性一致。血液学毒性是最常见的不良事件,包括≥3级贫血(60.9%)、血小板减少(47.8%)和中性粒细胞减少(47.8%)。恶心和腹泻等胃肠道不良事件很常见,但通常级别较低。值得注意的是,胃肠道毒性似乎比其他MDM2抑制剂报告的要轻。剂量限制性毒性主要发生在40mg队列中,因此明确将28天周期第1-5天每日一次30mg确定为推荐的2期剂量。尽管存在预期的血细胞减少,但联合用药的总体安全性是可控的,并且未观察到siremadlin暴露量增加与不良事件频率或严重程度之间的相关性。
在临床活性方面,芦可替尼-siremadlin联合用药成为该平台内评估的最活跃方案。在第24周,30.4%的患者实现了脾脏体积减少至少35%(SVR35),在30mg siremadlin队列中观察到显著的60%的反应率——这是ADORE所有组中最高的比例。中位脾脏体积减少总体达到-22.8%,在30mg队列中达到-38.7%,代表在已经接受芦可替尼治疗且反应不足的人群中,获得了有意义的额外益处。症状改善与这些解剖应答平行:21.7%的患者在第24周症状总评分 (TSS50) 降低≥50%,包括所有剂量队列的个体。鉴于长期芦可替尼暴露和基线时通常适度的症状负担,这些症状和体积反应尤其令人信服。
转化终点进一步支持了siremadlin的生物学活性。JAK2V617F等位基因负荷降低,第24周中位减少-12.6%,部分患者(包括伴有脾脏反应的患者)下降更深。SEALR突变患者也表现出变异等位基因频率的降低。这些发现与已知的长期芦可替尼单药治疗相关的典型缓慢且 中度的等位基因负荷减少相一致,并表明MDM2 抑制剂可能导致分子反应的增量获益。通过增加的循环GDF-15浓度,在所有siremadlin剂量水平上均证明了p53通路参与的药效动力学确认,在30mg队列中观察到最强的诱导。
一个重要的探索性观察结果为,在6名接受siremadlin治疗的患者中,出现了低变异等位基因频率的TP53突变(2-8%)。这些突变位于DNA结合域,符合功能丧失性变化,与其他MDM2抑制剂(如idasanutlin在PV中)的研究结果相似。尽管由于这些突变丰度低且缺乏详细的单细胞分析,其临床意义尚不确定,但它们强调了对克隆动力学进行仔细未来评估的必要性,特别是考虑到TP53改变在髓系肿瘤中已知的预后意义。治疗期间还在其他基因中检测到了其他突变,但其意义尚无法完全确定。
尽管研究入组的提前终止限制了平台设计的完全实现(特别是向第2和第3部分的过渡),但现有数据清楚地说明了ADORE方法的可行性和科学价值。在这种适应性平台的背景下,芦可替尼和siremadlin的联合治疗,特别是在30mg剂量水平上,显示出最一致和最令人信服的临床获益证据,并得到了分子和药效动力学活性以及可管理的毒性特征的支持。
综上所述,ADORE的结果将siremadlin定位为MF中芦可替尼的一个特别有前景的合作伙伴。该联合治疗不仅改善了JAK抑制后已确定存在次优反应的患者的脾脏大小和症状,还通过降低驱动基因突变负荷和靶向性激活p53信号,提供了疾病修饰的初步信号。尽管研究提前终止,但siremadlin在MPN中的未开发潜力仍然巨大。未来的研究可以探索其在更早期疾病阶段的作用,与其他新兴的疾病修饰疗法联合或序贯使用,并通过更深入的基因组表征来阐明其对克隆进化的影响。因此,siremadlin代表了MF及更广泛MPN谱系下一代治疗策略的重要候选药物,值得继续进行严格的临床探索。
BOREAS:MDM2抑制的III期验证
BOREAS试验是MDM2抑制剂在MF中的最终评估,比较了navtamadlin单药治疗与最佳可用疗法在JAK抑制剂治疗后复发/难治性疾病患者中的疗效。这项随机、多中心、全球性III期研究招募了183名TP53野生型MF患者,这些患者对先前的JAK抑制剂治疗难治或已复发(表1)。患者按2:1的比例随机分配接受navtamadlin 240mg,在28天周期中连续7天给药(随后21天停药期),或接受BAT,包括单药或联合治疗:羟基脲、化疗、免疫调节剂和支持性治疗;JAK抑制剂被排除在外。前两个周期给予抗腹泻预防用药,并在每个周期的navtamadlin治疗7天内给予抗恶心预防用药。主要终点为第24周的SVR35,次要终点包括TSS50、总生存期和无进展生存期。Navtamadlin安全且耐受性良好。最常见的3/4级治疗中出现的不良事件是血小板减少(37% vs 21%)、贫血(29% vs 25%)和中性粒细胞减少(28% vs 11%)。navtemadlin vs BAT 组胃肠道3/4级不良事件分别为腹泻 (5%vs 2%)、恶心 (3%vs 0%) 和呕吐 (2%vs 0%)。
在这项研究中,navtemadlin 在对既往 JAK 抑制剂治疗难治或复发的患者中展现了具有临床意义的疗效。在第 24 周,navtemadlin 组的脾脏体积减少 ≥35%(SVR35)和总症状评分改善 ≥50%(TSS50)的比例分别是 BAT(最佳可用疗法)组的三倍 [15% (18/123) vs 5% (3/60)] 和 两倍 [24% (30/123) vs 12% (7/60)]。
值得注意的是,对于既往 JAK 抑制剂治疗难治或复发的患者,navtemadlin 具有改善疾病负担生物标志物的潜力。这提示其具有抗克隆活性和疾病修饰作用。在 BOREAS 研究中,navtemadlin 的靶向生物学活性通过以下指标的显著下降得以体现:外周血 CD34+ 细胞计数、骨髓纤维化驱动基因的变异等位基因频率(VAF)降低 ≥50%,以及血清炎性细胞因子(如 TNF-α、IL-6 和 CRP)水平的下降。此外,navtemadlin 治疗引起的这些疾病生物标志物的变化与脾脏体积缩小的程度呈显著相关,证明了 navtemadlin 诱导的疾病修饰与治疗反应之间存在关联。最后,在拥有基线和第 24 周配对骨髓活检的 navtemadlin 治疗患者中,47% (31/66) 的患者报告骨髓纤维化分级改善了一级或以上。与先前在既往 JAK 抑制剂治疗难治或复发的患者中开展的 navtemadlin 单药治疗 II 期试验结果相似,III 期 BOREAS 研究的转化研究发现表明,通过治疗性干预改变驱动骨髓纤维化的潜在生物学机制,可以转化为切实的临床获益。
综上所述,生物标志物分析揭示了令人鼓舞的信号,表明 navtemadlin 可能靶向导致疾病的细胞并改变疾病进程。尽管详细的疗效结果有待最终发表,但在第 66 届美国血液学会年会上公布的数据支持了 MDM2 抑制在骨髓纤维化中的疾病修饰潜力。
近期临床经验带来的机制性见解
耐药机制与克隆演化
近期的临床经验为理解 MDM2 抑制剂的耐药机制提供了重要见解。一个令人担忧的观察结果是,在 MDM2 抑制剂治疗后出现了 TP53 突变的克隆。在一份病例报告中,一名 MDM2 扩增的腺样囊性癌患者在接受 MDM2 抑制剂治疗后,出现了持续的全面血细胞减少,并在外周血和骨髓中检测到 20 种不同的致病性 TP53 突变。这种现象被称为“MDM2 抑制剂相关性克隆性造血”,是一种以前未被认识的毒性反应,可能带来后续发生髓系恶性肿瘤的风险。在多名患者中检测到了血浆 TP53 突变,且突变数量与治疗持续时间呈强相关性。这些发现强调了对接受 MDM2 抑制剂治疗的患者进行仔细监测的必要性,并可能影响未来的临床试验设计。
实验室研究已证实,MDM2 抑制可在体外加速 TP53 突变克隆的选择。将 AML 细胞与递增浓度的 idasanutlin 长期培养,会导致携带 TP53 突变的耐药克隆出现。对 MPN 患者样本的单细胞分析进一步细化了这一演化模型,揭示 MDM2 抑制并非仅仅诱导突变发生,而是对预先存在的、罕见的 TP53 突变亚克隆施加了强大的选择性压力。在这些竞争性实验中,TP53 突变细胞表现出比野生型对应细胞更明显的适应优势,在治疗压力下迅速主导克隆结构。这种“克隆清除”效应表明,MDM2 抑制剂的暴露持续时间是一个关键变量,因为对 TP53 野生型主克隆的长期抑制为耐药性小亚克隆的扩增创造了一个允许性的生态位。值得注意的是,来自 siremadlin 和 navtemadlin 试验的长期随访数据一致表明,这种 TP53 突变小亚克隆的扩增在停药后通常是可逆的,并且截至目前,尚未明确证实其与继发性白血病发生率的增加有关。
除了遗传演化,还发现了一些独立于 TP53 状态的适应性耐药机制。抗凋亡 BCL-2 家族成员(尤其是 MCL-1)的上调,是对 p53 激活的一种代偿性生存通路反应。从机制上讲,MDM2 抑制可触发一个涉及 MCL-1 稳定的反馈回路,从而提高凋亡阈值,限制单药治疗的细胞毒性疗效。此外,最近的数据描述了一种由 CEBPB/IL-1β/TNF-α 反馈回路驱动的炎症性耐药轴,该轴存在于单核细胞性 AML 亚型中,即使在 p53 被强烈激活的情况下,也能促进白血病细胞存活。这些机制性见解对于理解 MDM2 抑制剂的长期安全性具有重要意义,并为联合策略(例如同时抑制 BCL-2 或 MCL-1)提供了强有力的理论依据,以减轻遗传性和非遗传性耐药的发生。
药效学生物标志物与患者选择
MDM2抑制剂的临床开发,与识别和验证能够可靠反映p53通路靶向结合、并实现合理患者选择的稳健药效学生物标志物紧密相关。在这些标志物中,循环中的生长分化因子15(GDF-15),也称为巨噬细胞抑制细胞因子-1(MIC-1),已成为一种特别有价值且实用的生物标志物。GDF-15是p53的直接转录靶点,在MDM2抑制后,其快速且剂量依赖性的诱导升高,提供了在体内功能性p53再激活的有力证据。在第一代MDM2拮抗剂RG7112的早期临床研究中, consistently 观察到血清GDF-15水平升高,并且与全身药物暴露相关,证实了在血液系统恶性肿瘤中的靶点结合。随后,在包括idasanutlin和siremadlin在内的下一代MDM2抑制剂中,也观察到了类似的药效学特征,从而使GDF-15诱导成为MDM2拮抗作用的一个类别定义性生物标志物。
除了可溶性生物标志物,对细胞内p53下游效应分子的调节也提供了更深入的机制解析。在MDM2抑制剂暴露后,已在白血病原始细胞和CD34+祖细胞群中记录到参与细胞周期阻滞(如CDKN1A/p21)、凋亡(如BBC3/PUMA、BAX)和应激反应的典型p53靶基因的转录上调。在RG7112的I期研究中,p53靶基因表达的诱导和p53蛋白的积累仅限于TP53野生型细胞,强调了这种治疗方法的生物学特异性。补充分析显示,凋亡标志物(包括Annexin V阳性和促凋亡蛋白的裂解)在早期即增加,这支持其作为治疗反应的早期指标,可在明显的临床获益出现之前发挥作用。
因此,MDM2抑制剂试验的患者选择策略已演变为一个以保留功能性p53信号为中心的、生物标志物驱动的框架。在各种肿瘤类型中,TP53野生型状态已成为一个基础的入组标准,反映了对完整p53转录活性的机制要求。重要的是,整合的基因组分析通过证明TP53突变与MDM2拷贝数改变之间的相互排斥关系,进一步细化了这一范式。在AML和MPN中,TP53突变在存在MDM2过表达或扩增的肿瘤中发生率显著较低,提示存在替代性的、相互排斥的p53通路抑制机制。这种相互排斥性已直接影响了后续临床试验的入组标准,富集了最有可能从MDM2抑制中获益的患者群体。这些选择原则的相关性在慢性髓系肿瘤中得到了进一步证实。在接受idasanutlin治疗的PV和ET患者中,p53通路的药效学激活(通过GDF-15诱导和恶性CD34+干细胞及祖细胞的耗竭证明)主要在TP53野生型疾病中观察到,且临床反应伴随着分子缓解和驱动基因突变等位基因负荷的降低。同样,在接受navtemadlin治疗的患者中,外周血CD34+细胞计数、MF驱动基因VAF和炎性细胞因子水平的降低转化为可证明的临床获益。这些发现支持了以下概念:药效学生物标志物不仅可确认靶点结合,还可能作为疾病修饰活性的替代指标。最近的联合研究将这些见解扩展到晚期MF。在ADORE平台试验中,siremadlin联合芦可替尼治疗导致循环GDF15水平持续升高,证实了联合治疗中p53通路的持续激活,同时在有反应的患者中观察到脾脏体积和JAK2V617F等位基因负荷的降低。
这些结果直接影响了正在进行的 3 期 POIESIS 试验(NCT06479135)的设计,该试验前瞻性地将入组限制为 TP53 野生型患者,并整合了生物标志物驱动的终点,以分离添加 navtamadlin 的临床贡献。总之,这些药效学和基因组学见解说明了生物标志物指导的开发如何将 MDM2 抑制从一种广泛应用的细胞毒性策略转变为一种精准医学方法。稳健的药效学生物标志物的开发对于优化 MDM2 抑制剂治疗至关重要。通过检测 GDF-15 水平升高来证明靶向性 p53 通路激活,为确认药物活性提供了一个实用的生物标志物。此外,促凋亡蛋白和细胞周期标志物的变化可以作为治疗反应的指标。
患者选择策略已发展到关注存在 MDM2 过表达或扩增证据的 TP53 野生型肿瘤。证明 TP53 突变随 MDM2 拷贝数增加而减少的相互排斥性分析,为当前临床试验的入组标准提供了信息。这些生物标志物驱动的方法代表了 MDM2 抑制剂向精准医学应用迈进的关键进展。
安全性考量与毒性管理
血液学毒性特征
血液学毒性是 MDM2 抑制剂的主要剂量限制性不良效应,在所有靶向 p53 再激活的临床项目中均持续观察到。这种类别效应反映了 MDM2 在正常造血干细胞和祖细胞内抑制 p53 活性的重要生理作用,在这些细胞中,不受控制的 p53 激活会导致细胞周期阻滞和凋亡。在早期和后期临床试验中,血小板减少、中性粒细胞减少和贫血是最常见的 3-4 级毒性,通常需要中断、减少剂量或停止治疗。其中,血小板减少是最主要且最早出现的剂量限制性毒性,通常发生在前一到两个治疗周期内,并与累积药物暴露密切相关。
对多个 MDM2 抑制剂试验(涵盖不同化学骨架和肿瘤适应症)的比较分析表明,骨髓抑制的模式高度一致,表明血液学毒性主要是由机制驱动而非化合物特异性。重要的是,这些观察结果直接塑造了现代给药范式。临床前和临床药代动力学-药效动力学模型显示,短暂的 p53 激活足以诱导恶性细胞凋亡,而正常造血细胞在停药后恢复更快。这种差异恢复动力学为间歇性给药策略提供了理论基础,该策略通过治疗间歇期让骨髓得以恢复,同时保留抗肿瘤疗效。
临床上,间歇性方案(例如,在 28 天周期内每周、每两周或短期给药)在未影响药效学靶点结合的情况下,显著提高了耐受性。这些进展使得下一代 MDM2 抑制剂的持续开发成为可能,并促进了联合策略的形成,特别是在血液系统恶性肿瘤中,平衡疗效和骨髓毒性仍然至关重要。
胃肠道及其他非血液学毒性
除了血液学效应外,MDM2 抑制剂始终会诱导胃肠道毒性,最常见的是恶心、呕吐、腹泻和食欲减退,这些是在早期和后期临床试验中报告的主要非血液学不良事件。这些毒性通常具有剂量和方案依赖性,是 p53 在快速增殖的胃肠上皮细胞中再激活的靶向效应,其中 p53 在调节细胞周期阻滞和凋亡中起着核心作用。在早期的基于 nutlin 的化合物(如 RG7112)中,胃肠道毒性常常限制了剂量递增和连续给药;第二和第三代抑制剂受益于改善的药代动力学和间歇性给药方案,这些方案显著减轻了严重程度。
预防性止吐方案、止泻剂和短期给药已成为标准的支持策略,并使得后期试验中能够持续给药。其他非血液学毒性包括疲劳、乏力、厌食以及偶尔出现的皮肤表现(如皮疹或瘙痒),这些通常是低级别的,并且可以通过中断治疗或调整剂量而逆转。重要的是,全面的心脏安全性评估已基本排除了新型 MDM2 抑制剂具有临床意义的 QT 延长,缓解了人们对长期给药的早期担忧。总体而言,尽管非血液学毒性仍然是一个定义性的类别效应,但通过对化合物选择性、给药方案和支持性治疗的持续优化,安全性特征已变得越来越可控,支持 MDM2 靶向治疗的持续临床开发。
未来方向与联合策略
合理的联合方法
MDM2 抑制剂开发的未来在于合理的联合策略,这些策略可以在管理毒性的同时增强疗效。芦可替尼联合 siremadlin 在 ADORE 中的成功为开发其他协同联合方案提供了模板。潜在的联合伙伴包括 JAK 抑制剂、BCL-2 抑制剂、CDK4/6 抑制剂和免疫检查点抑制剂。将 MDM2 抑制剂与其他靶向药物联合具有强大的生物学依据。MDM2 抑制会激活 p53 依赖性凋亡通路,这可以通过 BCL-2 抑制来增强。同样,MDM2 抑制的细胞周期效应可能与 CDK4/6 抑制剂协同。这些联合方案需要仔细优化剂量以平衡疗效和毒性。
JAK 抑制剂联合方案
在非临床研究中,Clevenger 等人表明,navtamadlin 和芦可替尼通过抑制 p21 介导的细胞周期阻滞,协同增强了 MF 患者来源的 CD34+ 祖细胞的凋亡。与非临床结果一致,navtamadlin 添加至芦可替尼治疗中对芦可替尼反应欠佳的 TP53 野生型 MF 患者的 1b/2 期 KRT-232-109 试验的临床数据,证明了 navtamadlin 添加至芦可替尼的 robust 疗效和协同作用,在第 24 周有 32% 的患者达到 SVR35。值得注意的是,患者在进入研究前已长期接受芦可替尼治疗,大多数服用 ≥15mg 每日两次的剂量,但仍无法达到最佳反应。不允许增加芦可替尼剂量,这证实了观察到的临床反应归因于 navtamadlin。这项 1b/2 期研究的结果支持了 III 期 POIESIS 试验的设计。
全球注册性 POIESIS 研究(NCT06479135)采用了一个结构严谨且临床实用的试验设计,以评估 navtamadlin 作为芦可替尼的添加疗法,用于对一线 JAK 抑制未能达到最佳反应的 JAK 抑制剂初治 MF 患者。POIESIS 设计的核心是一个长期芦可替尼单药治疗的导入期,该阶段反映了现实世界的治疗实践:在考虑额外治疗之前,启动、滴定并稳定芦可替尼治疗。大约 600 名患者将单独接受芦可替尼治疗 18-24 周,以便在标准化暴露下识别出不同的临床反应表型。达到最佳脾脏或症状反应的患者,以及原发性芦可替尼难治或疾病进展的患者将被终止入组,从而富集随机化人群,使其包括尽管芦可替尼剂量稳定但仍有明确次优反应的个体。这种方法最大限度地减少了异质性,并分离出了一个具有持续未满足临床需求的人群。随机化的资格还要求确认 TP53 野生型状态,从而确保保留 navtamadlin 对完整 p53 信号的机制依赖性。通过仅将次优反应者随机分配至 navtamadlin 或安慰剂组(均继续接受芦可替尼背景治疗),POIESIS 能够独特地分离出 MDM2 抑制的增量临床贡献,同时保持脾脏体积减少、症状改善以及长期结局(包括无进展生存期和总生存期)等既定终点的完整性。
免疫治疗联合方案
MDM2 抑制剂与免疫治疗药物的联合已成为一种潜在的治疗策略,临床前证据表明,MDM2 抑制可通过多种机制增强抗肿瘤免疫,包括上调 IL-15 和 MHC II 类产生以及改善肿瘤模型中的 T 细胞浸润。研究最深入的联合方案涉及免疫检查点抑制剂,特别是 PD-1/PD-L1 阻断剂。MDM2/p53 拮抗剂 alirozamdin 联合帕博利珠单抗的临床评估已在晚期、免疫治疗难治性实体瘤(包括黑色素瘤、非小细胞肺癌和尿路上皮癌)患者中显示出初步活性,在多个队列中观察到客观反应。另一种有效的 MDM2-p53 拮抗剂 brigimadlin 已在晚期 MDM2 扩增的胆道癌患者中显示出抗肿瘤活性,在一系列治疗患者中观察到部分反应,支持了正在进行的使用或不使用检查点阻断的 MDM2 抑制研究。一项正在进行的 I/II 期研究正在评估 milademetan 联合 atezolizumab,以更好地确定其在实体瘤中的耐受性和临床活性。
MDM2 抑制剂与干扰素的联合在 MPN 中显示出特别的前景,其中 RG7112 联合聚乙二醇化干扰素-α2a 在临床前异种移植模型中优先靶向 JAK2V617F 阳性的造血祖细胞并降低了突变等位基因 burden。基于 MDM2 抑制剂调节癌细胞免疫原性和影响 NKG2D 配体表达的能力,人们还探索了其与 CAR-T 细胞疗法的联合,这可能增强 CAR-T 的疗效,尽管临床证据尚未发表。在间皮瘤模型中,MDM2 抑制剂与溶瘤腺病毒的联合显示出协同细胞毒性,MDM2 抑制剂促进了病毒后代产生并放大了 p53 介导的凋亡通路。一种基于疫苗的方法,将 MDM2 衍生肽与 Nutlin-3 联合使用,显示肽特异性的 CD4+ T 细胞可通过颗粒酶 B 杀死肿瘤细胞,而 MDM2 抑制通过 CIITA 上调增强了这种反应。像 ALRN-6924 这样的双 MDM2/MDMX 抑制剂已在临床前和早期临床研究中显示出免疫调节特性,并在与检查点阻断联用时增加了 T 细胞浸润。
值得注意的是,MDM2 扩增已被确定为某些肿瘤类型中检查点抑制剂治疗期间出现超进展的一种机制,这表明将 MDM2 抑制与免疫治疗相结合可能减轻 MDM2 扩增情况下的肿瘤加速生长。临床开发一直受到预测性生物标志物的指导,其中野生型 TP53 是活性所必需的,而高水平的 MDM2 扩增与 MDM2 靶向治疗试验中增强的反应相关。尽管初步数据令人鼓舞,但挑战仍然存在,包括需要间歇性给药的靶向性血液学毒性,以及尽管有良好的 I 期活性,但一些研究性药物仍被中止。当前正在进行的试验继续在多瘤种中优化基于 MDM2 抑制剂的免疫治疗方案的联合方案、给药策略和患者选择标准。
下一代 MDM2 抑制剂与 PROTACs
“第一波”MDM2-p53 拮抗剂的主要局限性在于它们通过与 p53 结合口袋的占用发挥作用,并且矛盾的是,它们会加剧内在的 p53-MDM2 转录反馈环路:p53 激活诱导 MDM2 表达,增加了必须被饱和以维持通路再激活的抑制剂“汇”。蛋白质降解剂通过强制进行急性的、类似敲除的 MDM2 去除来重新构建这种药理学特性,从而抑制了反馈驱动的 MDM2 反弹,并将系统从慢性靶点覆盖转变为“打了就跑”的细胞命运承诺(细胞周期阻滞 vs 凋亡)。在 AML 相关背景中,招募 VHL 的 MDM2 降解剂 MS3227 体现了这一原理:它是从 AMG-232 衍生的 MDM2 配体改造而来,并通过连接体结构-活性关系进行了优化,强调了三元复合物几何结构和有效的泛素化(而非仅结合亲和力)决定了降解剂的性能。
机制上,MS3227 产生了快速、依赖于蛋白酶体和 cullin-E3 活性的 MDM2 耗竭,并表现出标志性的 PROTAC 行为,包括在高浓度下出现与无效二元复合物形成一致的钩状效应。在 TP53 野生型白血病细胞系中,MS3227 驱动的 p53 通路输出比匹配的化学计量抑制剂(AMG-232)更强,对典型 p53 靶点的诱导更显著,生长抑制作用更大,并且在洗脱后具有更持久的药效学影响——这些特征与 MDM2-p53 回路中催化性降解和改变的反馈动力学相一致。对于转化定位而言重要的是,MS3227 的活性扩展到了基质共培养中的原代 AML 标本,在特定样本中,它更倾向于减少白血病原始细胞群而非更不成熟的 CD34+ 区室,并且在 TP53 缺失的疾病中活性有限——这再次强调了,尽管 MDM2 可以不受 TP53 基因型影响而被生化降解,但治疗依赖性仍然强烈地富集于 TP53 野生型背景中。
迄今为止,该领域最先进的临床前数据集可能由 KT-253 提供(表1),这是一种高效力、招募 CRBN 的异双功能 MDM2 降解剂,旨在通过阻止急性 MDM2 反馈蓄积和实现间歇性给药来表现优于小分子抑制剂。在细胞分析中,KT-253 达到了亚纳摩尔的 MDM2 降解效力,并将其转化为跨血液学模型的皮摩尔至亚纳摩尔的功能活性,而对照的小分子抑制剂则诱导了 MDM2 上调而不是耗竭,并且在匹配的分析中效力低数个数量级。这种效力与快速的通路参与相关联:KT-253 以非常高的表观效率稳定 p53,并诱导广泛的转录性 p53 程序,其浓度远低于 DS-3032 仅产生中度效应的浓度。一个关键的概念性进展是证明了短暂暴露足以产生不可逆的凋亡承诺:短暂的 KT-253 处理随后洗脱,仍在 24-48 小时内产生 caspase 激活,支持了可能扩大治疗窗口的间歇性方案。在体内,这种“脉冲药理学”转化为显著的异种移植疗效。在 MV4;11 AML 异种移植模型中,单次静脉注射 KT-253 在 6 只动物中的 5 只中产生了完全缓解;而临床相关的 DS-3032 方案在该头对头研究中没有产生完全缓解。除了肿瘤体积终点外,该报告还显示,与暴露匹配的小分子抑制剂给药相比,KT-253 暴露后出现了中度的 p53 通路激活和凋亡执行(cleaved caspase-3)。临床上相关的联合逻辑也正在出现:KT-253 在 AML 模型中与维奈克拉协同,在 MOLM-13 异种移植模型中,单次 KT-253 剂量与维奈克拉方案联合产生了持久的完全缓解,而任何一种药物单独在该耐药模型背景下均未能达到 CR。与此轴一致,MS3227 研究同样观察到了与 MCL-1 下调相关的维奈克拉协同作用,这是克服 AML 中常见维奈克拉耐药节点的机制基础。
从开发角度来看,这些项目还阐明了 MDM2 降解剂的核心设计权衡:(i) E3 连接酶选择(CRBN vs VHL)以平衡肿瘤表达模式、降解深度和耐受性负担;(ii) 连接体工程以优化有效的三元复合物形成;以及 (iii) 选择性和新底物风险,特别是对于 IMiD 样 CRBN 招募剂,其中脱靶新底物降解是一个类别关注点(尽管 KT-253 在 RS4;11 中的深度蛋白质组学分析显示出高度表观选择性,p53 通路上调主导了反应特征)。总之,临床前证据支持以下观点:下一代 MDM2 降解剂可能实现更持久的通路再激活,且不需要连续暴露,从而可能相比小分子抑制剂扩大治疗指数——尤其是在 TP53 野生型髓系恶性肿瘤中,短暂的、高强度的靶点耗竭后似乎可实现凋亡“承诺”。
最后,近期的会议报告强调了相同的区分主题——反馈环路抑制、相对于多种临床探索的小分子抑制剂的效力,以及间歇性给药策略和生物标志物选择的适应症(包括 AML)的理由。
结论
靶向 MDM2-p53 轴代表了现代精准医学中最具转化价值的成功案例之一,从基础结构生物学发现进展到在髓系恶性肿瘤中的临床验证。虽然早期 MDM2 抑制剂在 AML 中的开发面临着挑战性的毒性特征和 modest 的单药活性,但关键的经验教训——包括间歇性给药以管理血液学毒性、通过 TP53 突变状态和 MDM2 扩增进行生物标志物驱动的患者选择,以及通过炎症反馈环路产生耐药性的识别——共同塑造了当前的临床开发格局。
在 MPN 中,MDM2 抑制已从概念验证进展为有临床意义的疾病修饰活性,如 BOREAS 和 ADORE 试验所示,这些试验证明了脾脏、症状和分子学反应的改善。下一代 MDM2 降解剂和基于 PROTAC 的方法通过实现更深度的靶点耗竭和解除反馈驱动的 MDM2 上调,提供了改善治疗指数和克服小分子抑制剂局限性的潜力。随着该领域推进联合策略——包括 JAK 抑制剂、BCL-2 拮抗剂和免疫疗法——以及实施创新的适应性平台试验(如 ADORE 和 POIESIS),MDM2 抑制剂正在成为髓系恶性肿瘤(尤其是MPN)治疗工具箱中越来越重要的组成部分,为靶向 p53 通路在癌症治疗中提供了新的方向。
参考文献
Leukemia . 2026 May 5. doi: 10.1038/s41375-026-02975-6