深度解析医学证据,DeepEvidence为你支撑决策
背景介绍
胃炎是一种常见的胃肠道疾病,其发病机制传统上归因于幽门螺杆菌感染、药物刺激或自身免疫因素。然而,临床观察发现,即使在成功根除幽门螺杆菌后,部分患者仍会经历胃炎症状的复发,提示除了局部因素外,全身性调节机制也参与了疾病进程。近年来的神经免疫学进展强调了大脑-免疫双向沟通是理解炎症性疾病的关键框架。大脑通过感觉输入接收来自外周器官的免疫相关信号,并协调对免疫挑战的应答。然而,大脑是否以及如何编码和检索胃炎免疫信号,目前仍不清楚。下丘脑室旁核是神经内分泌和自主神经整合的中枢,在应激反应和内脏调节中发挥核心作用。本研究旨在揭示PVN中的印迹神经元如何编码胃部炎症信息,并探讨其在慢性炎症致敏化和应激诱导胃炎恶化中的作用机制。
研究思路
针对上述科学问题,山东大学基础医学院的孙晋浩(Jinhao Sun)教授团队开展了一项系统性研究。通过HCl/乙醇诱导的小鼠胃炎模型,结合活动依赖性遗传标记、化学遗传学和光遗传学技术,该团队发现胃部炎症信息可同时通过迷走神经(神经通路)和循环IL-1β/TNF-α(体液通路)传递至PVN,激活Fos阳性神经元(Fosᴾⱽᴺ)。这些神经元通过两条平行且协同的通路调节胃炎:下丘脑-垂体-肾上腺轴(CRH依赖)和PVN-迷走神经背核-胃神经回路。单核RNA测序揭示,胃炎状态下Fosᴾⱽᴺ神经元中Hcn1表达显著上调。抑制Hcn1可降低神经元兴奋性并减轻胃部病理。值得注意的是,反复激活Fosᴾⱽᴺ神经元可形成特异性的“炎症致敏化”,导致疾病持续存在;而束缚浸水应激和社会挫败应激可通过重新激活约80%的胃炎激活Fosᴾⱽᴺ印迹神经元来加剧胃部炎症。相关内容以Hcn1-dependent engram neurons in the PVN encode gastric inflammatory sensitization为题,发表在Science Advances!

图片解析
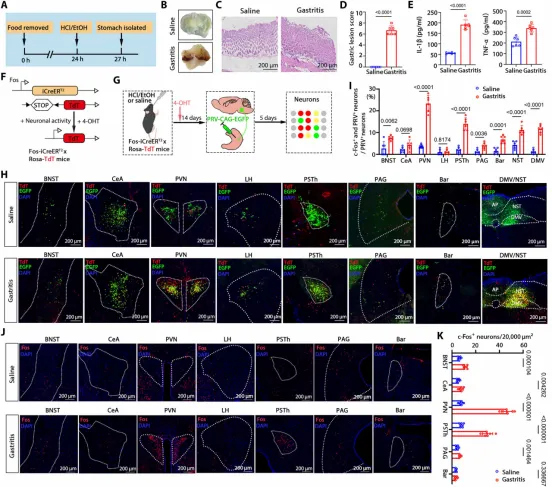
图1. 胃炎模型中Fosᴾⱽᴺ神经元被特异性激活: (a) HCl/EtOH诱导胃炎模型示意图。(b) 胃组织大体形态。(c,d) H&E染色及黏膜损伤评分。(e) 胃组织中IL-1β和TNF-α水平升高。(f) 4-OHT处理后激活神经元中TdTomato表达示意图。(g) PRV逆行示踪实验流程。(h) 全脑PRV-EGFP⁺神经元图谱。(i) 各脑区c-Fos/PRV-EGFP共定位定量,PVN共定位率最高。(j,k) PVN和PsTh中c-Fos⁺神经元计数。
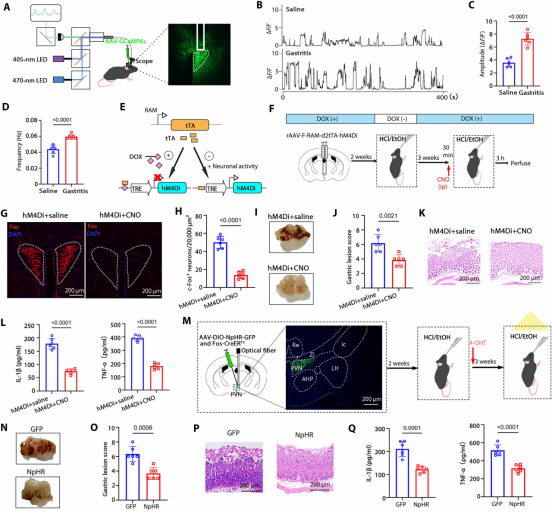
图2. Fosᴾⱽᴺ神经元调节胃炎: (a) 钙成像实验装置示意图。(b) PVN神经元代表性钙信号。(c,d) 胃炎组PVN神经元钙信号振幅和频率显著增加。(e) RAM活动标记系统示意图。(f) 化学遗传学抑制Fosᴾⱽᴺ神经元实验流程。(g,h) 化学遗传学抑制后PVN中Fos表达显著降低。(i-k) 胃损伤评分及H&E染色。(l) 化学遗传学抑制后胃部炎症细胞因子水平降低。(m) 光遗传学抑制Fosᴾⱽᴺ神经元实验流程。(n-p) 胃损伤评分及H&E染色。(q) 光遗传学抑制后炎症细胞因子水平降低。
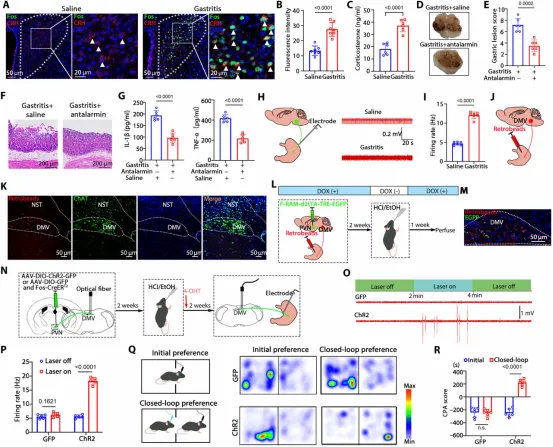
图3. Fosᴾⱽᴺ神经元通过HPA轴和Fosᴾⱽᴺ-DMV-胃回路调节胃炎: (a) PVN中Fos与CRH共定位免疫荧光。(b) 胃炎组CRH表达显著增加。(c) 胃炎组血清皮质酮水平升高。(d-g) CRH受体拮抗剂antalarmin预处理后胃损伤减轻。(h) 胃神经电活动记录示意图。(i) 胃炎组胃神经放电频率增加。(j) 胃壁注射红色逆行微球实验方案。(k) 红色逆行微球与ChAT神经元在DMV中共定位。(l) PVN注射F-RAM-d2tTA-TRE-EGFP病毒联合胃逆行微球注射实验时间线。(m) DMV中逆行微球与Fosᴾⱽᴺ轴突末梢共定位。(n) PVN光遗传学病毒注射联合DMV光纤植入实验设计。(o,p) 光刺激PVN末梢后胃神经放电频率显著增加。(q) CPA实验中运动轨迹热图。(r) 光刺激前后CPA评分。
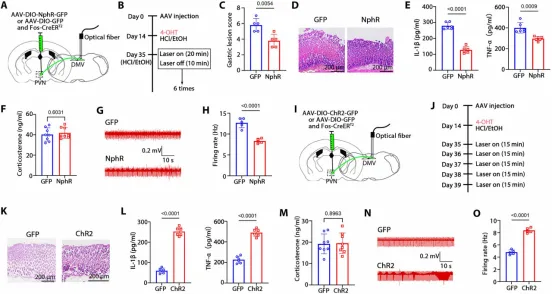
图4. Fosᴾⱽᴺ神经元通过Fosᴾⱽᴺ-DMV回路调节胃炎: (a,b) PVN-DMV回路抑制实验示意图。(c,d) 回路抑制后胃损伤评分及H&E染色。(e) 回路抑制后胃炎症细胞因子水平降低。(f) 回路抑制后血清皮质酮无显著变化。(g,h) 回路抑制后胃神经放电频率降低。(i) PVN-DMV回路激活实验示意图。(j) PVN-DMV回路激活实验设计。(k) 回路激活后H&E染色。(l) 回路激活后胃炎症细胞因子水平升高。(m) 回路激活后血清皮质酮无显著变化。(n,o) 回路激活后胃神经放电频率增加。
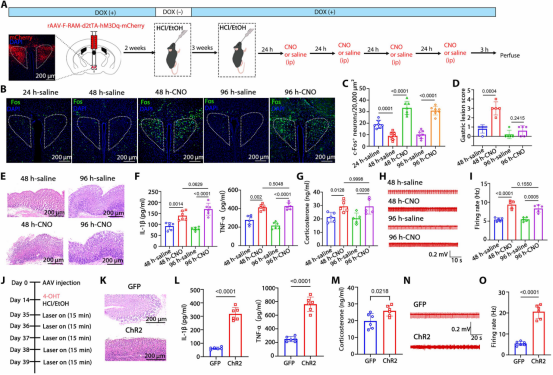
图5. 反复Fosᴾⱽᴺ激活建立慢性炎症: (a) 化学遗传学激活Fosᴾⱽᴺ神经元实验时间线。(b,c) 48和96小时时CNO组Fos免疫荧光显著高于对照组。(d,e) CNO组胃损伤评分和H&E染色显示胃壁结构紊乱。(f) CNO组胃炎症细胞因子水平显著升高。(g) CNO组血清皮质酮水平升高。(h,i) CNO组胃神经放电频率增加。(j) 胃炎完全消退后连续5天光遗传学激活Fosᴾⱽᴺ神经元实验设计。(k) 激活组H&E染色显示胃壁细胞紊乱。(l) 激活组胃炎症细胞因子水平显著升高。(m) 激活组血清皮质酮无显著变化。(n,o) 激活组胃神经放电频率增加。
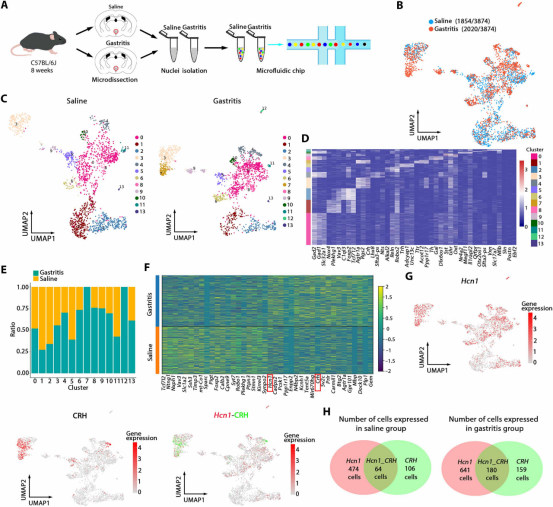
图6. 单核RNA测序鉴定Hcn1为Fosᴾⱽᴺ神经元的关键调节因子: (a) PVN snRNA-seq实验流程。(b) Fos神经元转录组UMAP图。(c,d) Fos神经元亚群聚类分析及标记基因热图。(e) 组间Fos神经元簇比例变化。(f) 差异表达基因热图,Hcn1和CRH显著上调。(g) Hcn1与CRH共表达UMAP可视化。(h) 胃炎组Hcn1⁺/CRH⁺细胞数量增加。
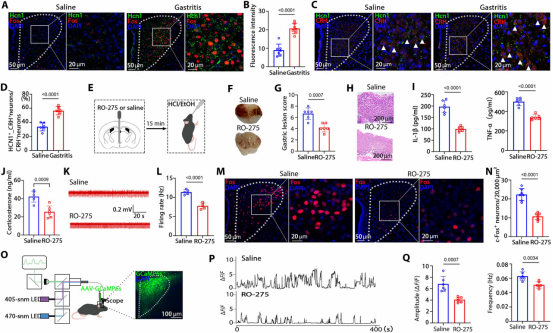
图7. Hcn1是Fosᴾⱽᴺ神经元的关键调节因子: (a) Hcn1与Fos双重免疫荧光代表图。(b) 胃炎组Hcn1表达上调。(c) Hcn1/CRH共染代表图。(d) 胃炎组Hcn1/CRH共定位比例升高。(e) PVN靶向注射Hcn1抑制剂RO-275示意图。(f-h) RO-275处理后胃损伤评分及H&E染色。(i) RO-275处理后胃炎症细胞因子水平显著降低。(j) RO-275处理后血清皮质酮水平降低。(k,l) RO-275处理后胃神经放电频率降低。(m) 生理盐水组与RO-275组Fos免疫荧光代表图。(n) RO-275组Fos表达降低。(o) PVN钙成像实验示意图。(p) 生理盐水组与RO-275组PVN神经元钙信号代表图。(q) RO-275处理后钙信号振幅和频率显著降低。
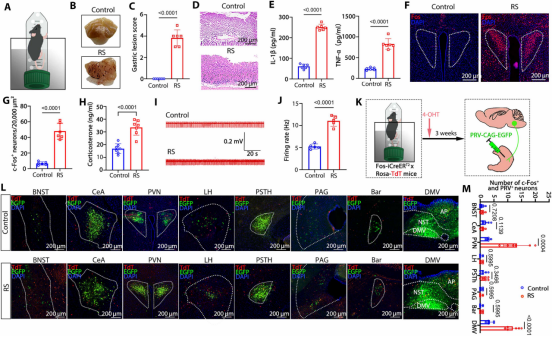
图8. 应激通过激活Fosᴾⱽᴺ神经元加剧胃炎: (a) 束缚浸水应激模型实验设计。(b,c) 应激组胃出血点和黏膜损伤评分。(d) 应激组胃壁细胞排列紊乱。(e) 应激组胃炎症细胞因子水平升高。(f,g) 应激组PVN中Fos⁺神经元增加。(h) 应激组血清皮质酮水平升高。(i,j) 应激组胃神经放电频率增加。(k) 水浸束缚应激期间标记Fos阳性神经元联合胃壁PRV注射实验设计。(l,m) 应激组Fosᴾⱽᴺ神经元向胃壁的投射最丰富。
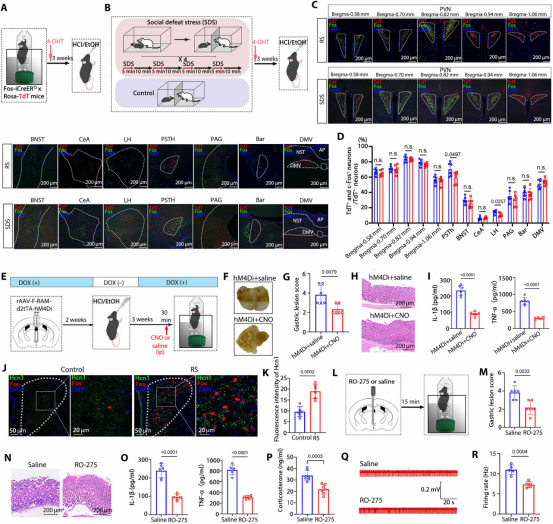
图9. 应激通过Fosᴾⱽᴺ神经元激活加剧胃炎: (a) 束缚应激与胃炎诱导Fos激活比较实验设计。(b) 社会挫败应激与胃炎诱导Fos激活比较实验设计。(c) 两种应激诱导的Fos神经元与胃炎诱导Fos神经元的共定位代表图。(d) 重叠率定量,两种应激均约80%。(e) 应激期间Fosᴾⱽᴺ神经元抑制实验设计。(f-h) 抑制后胃损伤评分及H&E染色。(i) 抑制后应激诱导的胃炎症细胞因子水平降低。(j) PVN中Fos与Hcn1双重免疫荧光代表图。(k) 应激组Hcn1表达升高。(l) PVN靶向注射RO-275实验方案。(m,n) RO-275处理后胃损伤评分及H&E染色。(o) RO-275处理后胃炎症细胞因子水平显著降低。(p) RO-275处理后血清皮质酮水平降低。(q,r) RO-275处理后胃神经放电频率恢复正常。
结论
本研究首次揭示了下丘脑室旁核中的Fos阳性印迹神经元编码并调节胃部炎症的机制。胃部炎症信息通过迷走神经(神经通路)和循环IL-1β/TNF-α(体液通路)双途径传递至PVN,激活Fosᴾⱽᴺ神经元。这些神经元通过HPA轴和PVN-迷走神经背核-胃神经回路两条平行通路调节胃部炎症。单核RNA测序鉴定出Hcn1是调控Fosᴾⱽᴺ神经元兴奋性和胃炎进展的关键分子。反复激活Fosᴾⱽᴺ神经元可形成“炎症致敏化”,驱动胃炎持续存在;而心理应激可通过重新激活约80%的胃炎激活Fosᴾⱽᴺ印迹神经元来加剧胃部炎症。该研究阐明了大脑存储和检索免疫信息的神经机制,并将Hcn1确定为慢性炎症性疾病神经调控治疗的潜在靶点。
原文链接:
https://www.science.org/doi/10.1126/sciadv.aeb6961