首页 > 健康资讯/ 正文
深度解析医学证据,lxfs.net为你支撑决策
你有没有在某个瞬间问过自己:我更容易被异性还是同性吸引,到底是自己的选择,还是我生来如此?
这个问题纠缠了人类很久。有人说性取向是家庭环境、成长经历塑造的印记;也有人坚信,这不过是出生时就已经写好的剧本。两种声音谁也说服不了谁,直到实验室里的基因测序仪开始介入这场争论。
1993年,美国科学家Dean Hamer在《科学》杂志上发表了一项研究:他对40对同性恋兄弟进行了基因分析,发现其中33对在X染色体的Xq28区域共享了相同的遗传标记。这是科学界第一次提出“可能存在同性恋基因”的证据,引发轩然大波——有人欢呼“性取向天生”终于有了科学背书,也有人质疑样本太小、无法重复。
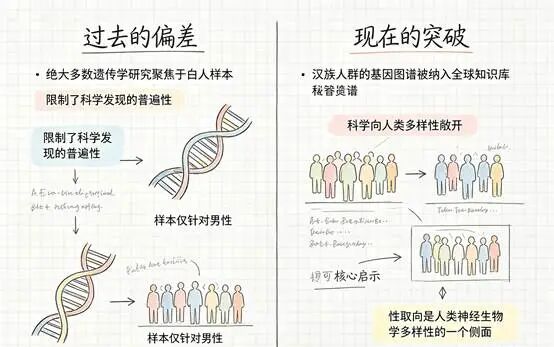
近三十年里,后续研究有的支持、有的否定,争议从未真正平息。而且绝大多数研究都聚焦于欧洲裔白人,一个关键问题始终悬而未决:这些发现换到完全不同的人群背景里,还成立吗?
2021年,浙江大学医学院附属第一医院和北京大学第六医院的研究团队在Cell Discovery上发表了一项针对中国汉族人群的大规模研究。他们没有找到所谓的“同性恋基因”,却发现了更复杂的东西。要理解这个答案,得先从最基础的层面开始——基因和性取向之间,到底有没有关系?
基因里,到底有没有“同性恋密码”
科学界寻找“与性取向有关的基因”,最常用的工具叫全基因组关联分析(GWAS)。它的逻辑很简单:把同性恋和异性恋的基因组放在一起,逐个位点比较,看哪些位置存在系统性的差异——不预设候选基因,让数据自己说话。
这项研究做了两阶段分析,总共纳入了1478名同性恋男性和3313名异性恋男性,全部为汉族人群。这是迄今为止针对中国人群规模最大的性取向遗传学研究。
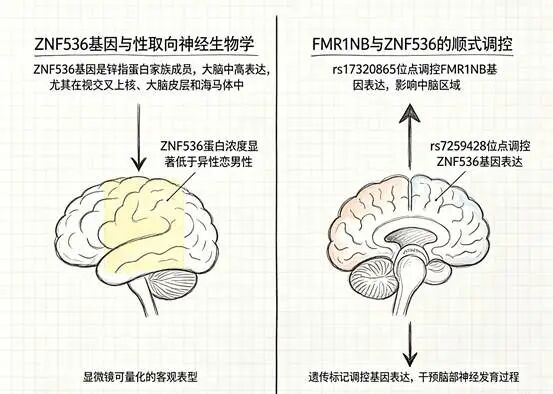
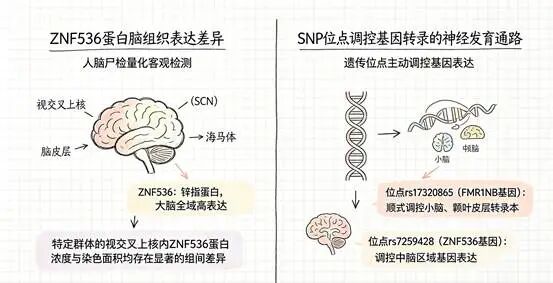
两个基因位点浮出了水面。第一个在X染色体上,叫rs17320865,藏在FMR1NB基因内部。X染色体本身就很引人注意——男性只有一条X染色体(来自母亲),如果上面有什么“决定性取向”的变异,表达起来不受另一条X的“对冲”影响。第二个在19号染色体上,叫rs7259428,位于ZNF536基因内部。
两个位点的P值都达到了全基因组显著性水平(P<5×10⁻⁸)——统计学上这是一个相当苛刻的标准,意味着它们不太可能是随机噪音。
比值比(OR)能告诉我们关联的强度。rs17320865的OR是1.31,意味着携带这个基因变异的个体,同性恋倾向的几率是未携带者的1.31倍;rs7259428的OR是0.65,意味着携带者几率降低约35%。这两个数字都不算大,这是否意味着,没有哪个基因变异能“决定”一个人是不是同性恋,它们只是在大样本统计中显现出的微弱信号?
当研究团队把中国汉族的数据与超过40万欧洲人群的基因组数据合并分析后,又发现了三个新的全基因组显著位点。跨种族的遗传重叠说明了一个重要事实:性取向的生物学基础可能具有超越特定人群的共性,它不是某种文化背景下独有的产物。
但人群遗传学研究有一个天然的局限——它只能告诉你“这个地方有关联”,却无法回答“这个关联到底怎么影响行为”。而且,把所有已知基因位点的信息加在一起,多基因风险评分也只能解释性取向差异中极小的一部分——大约1.11%到2.34%。基因有影响,但远不是“决定”。
大脑里有没有“同性恋痕迹”
基因位点找到了,却无法回答一个更根本的问题:这些基因到底做了什么,才让一个人的性取向发生偏移?
要回答这个问题,科学家必须离开人群统计数据、走进实验室。
FMR1NB基因的功能此前并不清晰,只知道它和脑发育、阅读能力可能沾边。为了弄清楚它在行为层面到底扮演什么角色,研究团队用CRISPR基因编辑技术敲除了小鼠的FMR1NB基因。
结果耐人寻味。和正常小鼠相比,敲除了FMR1NB的雄性小鼠对同性入侵者的爬跨行为明显增加,对雌性的偏好也变得模糊。更深层的发现在分子层面——这些小鼠的外周血转录组显示,多巴胺和血清素代谢通路发生了显著改变。
这两种神经递质你可能听说过,它们不直接“制造”性取向,但参与调控情感、奖赏和本能行为。想象一下:当你看到喜欢的人时,脑子里那种愉悦的感觉,很大程度上就是多巴胺在起作用。基因缺失扰动了这套系统,小鼠的行为随之变化。
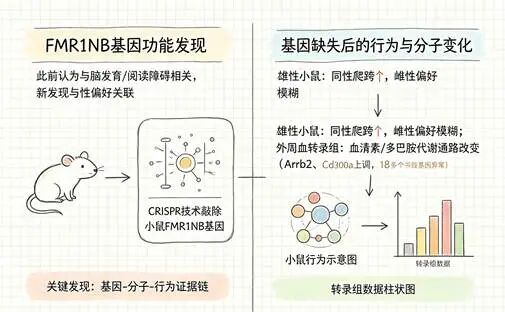
但必须诚实地说:小鼠的“爬跨”行为不能等同于人类的性取向。在啮齿类动物身上,爬跨同时涉及社会支配和性行为,概念维度完全不同。这份证据的性质是“提示性”而非“确定性”:这个基因参与调控社会行为,可能的路径是通过神经递质通路,但它不是“同性恋基因”。
更有力的证据来自人脑。研究团队从荷兰人脑库获取了26例去世后捐赠的脑组织标本——13例来自同性恋男性,13例来自异性恋男性。他们把目光投向一个叫做视交叉上核的脑区。这个区域只有米粒大小,位于下丘脑深处,是大脑昼夜节律的核心调控中枢。它之所以被选中,有一个重要的科学史背景:1990年,科学家就发现同性恋男性的视交叉上核体积大约是异性恋男性的两倍——而且这个脑区是目前已知唯一与性取向相关、却不单纯与性别相关的大脑结构。
这项研究往前又走了一步。研究人员在显微镜下定量分析了视交叉上核中ZNF536蛋白的浓度和覆盖面积,发现同性恋男性该蛋白浓度显著低于异性恋男性(0.011±0.001 vs 0.021±0.004,P=0.013),染色面积也更小(0.075±0.040 vs 0.137±0.103,P=0.043)。这不是问卷上的自我报告,也不是行为观察者的主观判断——是显微镜下可以量化的客观差异。
人群遗传学找到了关联位点,小鼠实验显示了行为改变,人脑组织测出了蛋白差异。三条证据线从不同层面指向同一个方向:性取向有生物学基础。但“有生物学基础”不等于“完全由基因决定”。大脑中那些与性取向相关的差异是真实的,可大脑本身也在成长中不断被经历塑造。
这自然引出了那个绕不开的问题:除了先天因素,后天经历到底扮演了什么样的角色?
基因不是全部,那剩下的是什么?
双胞胎研究给出过一个被反复引用的数字:男性性取向的遗传率约为60%。这意味着在群体层面,性取向的差异大约有六成可以归因于基因差异——但同时也意味着还有四成来自别处。
这个数字需要正确理解。遗传率描述的是某个特定样本里“基因能解释多少差异”,而不是“一个人的性取向有60%被基因决定”。不同研究的遗传率估算从30%到60%不等——差异本身就在提醒我们,这不是一个固定不变的常数。它更像一个快照,拍的是特定人群、特定时代下的情况。
如果换一个角度看,基因的解释力就更显有限了。前面提到的多基因风险评分,把所有已知的基因位点信息加在一起,也只能解释性取向差异中不到3%的方差。换句话说,即便你把自己的全基因组都测一遍,把所有与性取向相关的位点都找出来,它们加起来依然几乎无法预测你究竟会对谁产生爱意。
那剩下的部分是什么?可能是母亲子宫内的激素环境,可能是青春期的某些经历,也可能是成年后无法被基因测序仪捕捉到的各种生命经验。基因和环境从来不是各占一半、互不干扰的两个独立因素——它们始终在相互作用。一个基因变异可能在某种环境下影响显著,在另一种环境下则几乎不发挥作用。
把性取向简单拆解为“基因占多少、环境占多少”,本身就是一个过于简化的提问方式。但它指向一个更真实的答案:这不是二选一,而是交织。
当科学不再回答“是或否”
“同性恋是天生的还是后天的”——这个问题本身,或许就问错了方向。
它预设了一个前提:先天和后天是可以分开的,像水与油那样泾渭分明。但真实的生命不是这样运作的。基因写下的是草稿,不是终稿;大脑提供的是倾向,不是宿命。那个草稿在子宫里被激素修改,在童年被经历涂抹,在青春期被情感重写——到最后,你很难说清楚哪个笔画属于基因,哪个属于环境。因为它们本来就在一起。
三十年前,Hamer的发现让人们以为找到了“同性恋基因”;三十年后,中国人群的大规模研究告诉我们,基因确实有影响,但没有任何一个基因位点能决定一个人的性取向。
科学最诚实的贡献,或许不是给出一个干脆的“是或否”,而是把问题本身变得更审慎。
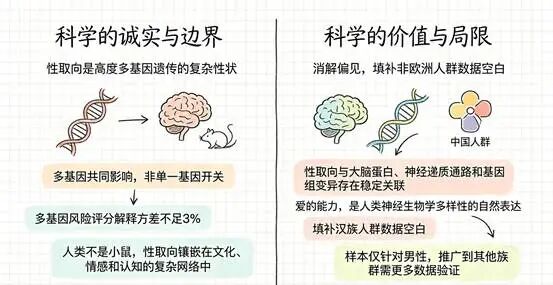
这或许就是科学最诚实的贡献。它存在的意义,从来都不是让世界变得更简单,而是让我们学会与复杂共处。它告诉我们:性取向既不是纯粹的选择,也不是纯粹的被决定——它是基因、发育、环境、经历交织在一起的产物,是人类神经生物学多样性的自然表达。
这个答案没有二元对立那么干脆,但它更接近真相。
参考资料
1. Hu S.et al. Cell Discov. 2021 Oct 31;7(1):103.
2. https://doi.org/10.1016/0006-8993(90)90350-K
小提示:本篇资讯仅在梅斯医学APP中开放阅读,请扫描二维码直接下载APP
