深度解析医学证据,DeepEvidence为你支撑决策
背景介绍
亚单位疫苗在诱导强效细胞免疫和交叉保护方面存在明显不足,难以应对癌症和不断突变的病毒(如SARS-CoV-2)。疫苗在体内的时空命运——从注射部位到淋巴结,再到抗原呈递细胞(APC),最终到达抗原加工的关键细胞器——是决定免疫应答质量和强度的核心。目前多数疫苗递送系统主要关注淋巴结积累和APC摄取,而忽略了将抗原精准递送至内质网(ER)——MHC I类分子合成与抗原肽加载的关键位点。因此,实现“淋巴结-APC-内质网”(LAE)级联靶向递送,是增强CD8⁺ T细胞介导的细胞免疫和交叉保护的有效策略。
研究思路
针对上述挑战,四川大学华西药学院孙逊教授团队提出了一种基于尖刺形貌介孔二氧化硅纳米颗粒的LAE级联递送策略。通过调控十六烷基三甲基溴化铵浓度和反应时间,合成了无刺(MN)、短刺(SNS)和长刺(SNL)三种形貌的介孔二氧化硅纳米颗粒,其中长刺纳米颗粒(SNL)在细胞摄取、DC成熟和抗原交叉呈递方面表现最优。机制研究表明,SNL通过机械刺激激活树突状细胞上的Piezo1离子通道,促进钙离子内流,进而激活钙调蛋白-钙调神经磷酸酶-NFAT信号通路,上调免疫相关基因表达。更重要的是,钙内流改变了囊泡运输路径,使SNL携带的抗原从传统的溶酶体逃逸转向优先递送至内质网,从而显著增强MHC I类分子介导的交叉呈递。利用这一ER靶向优势,团队将STING激动剂2'3'-cGAMP与抗原肽共包载于SNL中,实现了抗原与佐剂在ER水平的协同递送,进一步放大了免疫激活。在淋巴瘤(EG7-OVA)、宫颈癌(TC-1)和SARS-CoV-2疫苗模型中,SNL共载疫苗均诱导了强效的CD8⁺ T细胞应答,延缓肿瘤进展,并表现出对新冠病毒变异株的交叉保护能力。相关内容以“A Morphology‐Driven Cascade Delivery of Antigens for Potent T Cell Immunity”为题,发表在Advanced Materials。

图片解析
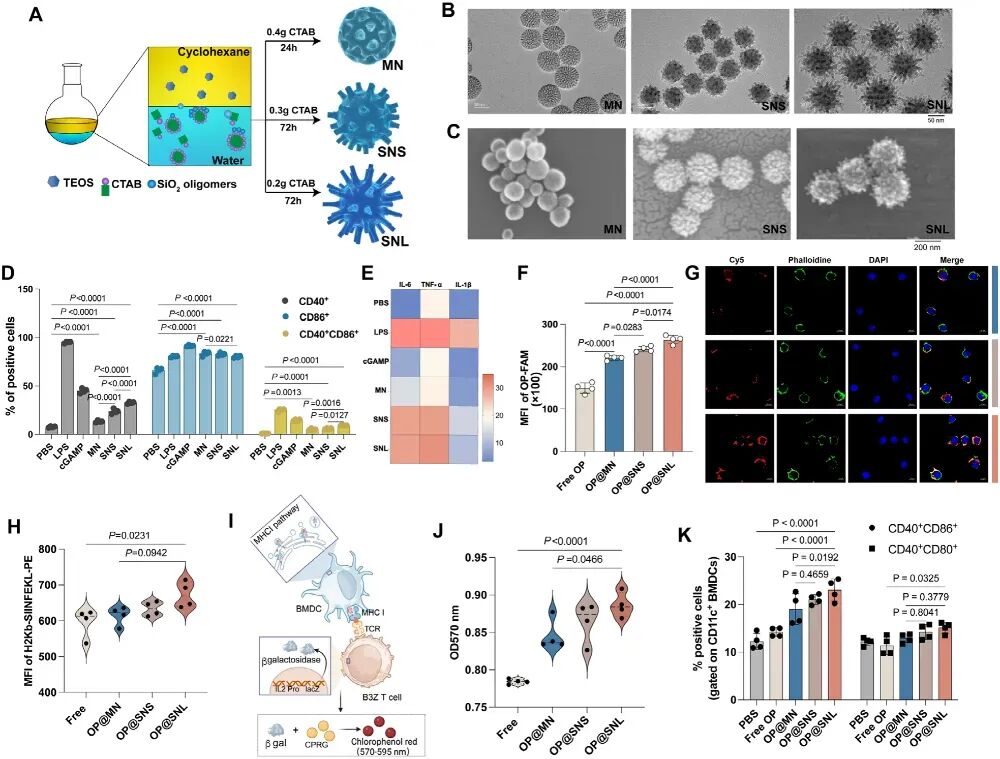
图1. SNL具有更高的细胞摄取效率和更强的细胞刺激作用: (A) 不同形貌纳米颗粒的合成示意图。(B) MN、SNS、SNL的透射电镜图像(比例尺50 nm)。(C) 扫描电镜图像(比例尺200 nm)。(D-E) BMDCs与纳米颗粒(20 μg/mL)孵育24 h后,CD40⁺、CD86⁺、CD40⁺CD86⁺细胞百分比及IL-6、TNF-α、IL-1β分泌水平(n=5)。(F) 流式细胞术检测负载FAM-OP的纳米颗粒在DC2.4细胞中的摄取效率(1 h,n=4)。(G) 激光共聚焦显微镜显示负载Cy5-OP的纳米颗粒在DC2.4细胞中的摄取(红色:Cy5-OP,绿色:细胞膜,蓝色:细胞核)。(H-J) 抗原交叉呈递和T细胞活化:BMDCs处理后检测H2Kb SIINFEKL-PE⁺细胞比例(H),与B3Z T细胞共培养后检测CPRG吸光度(I-J)(n=4)。(K) 不同纳米颗粒负载OP后诱导BMDCs成熟(n=4)。
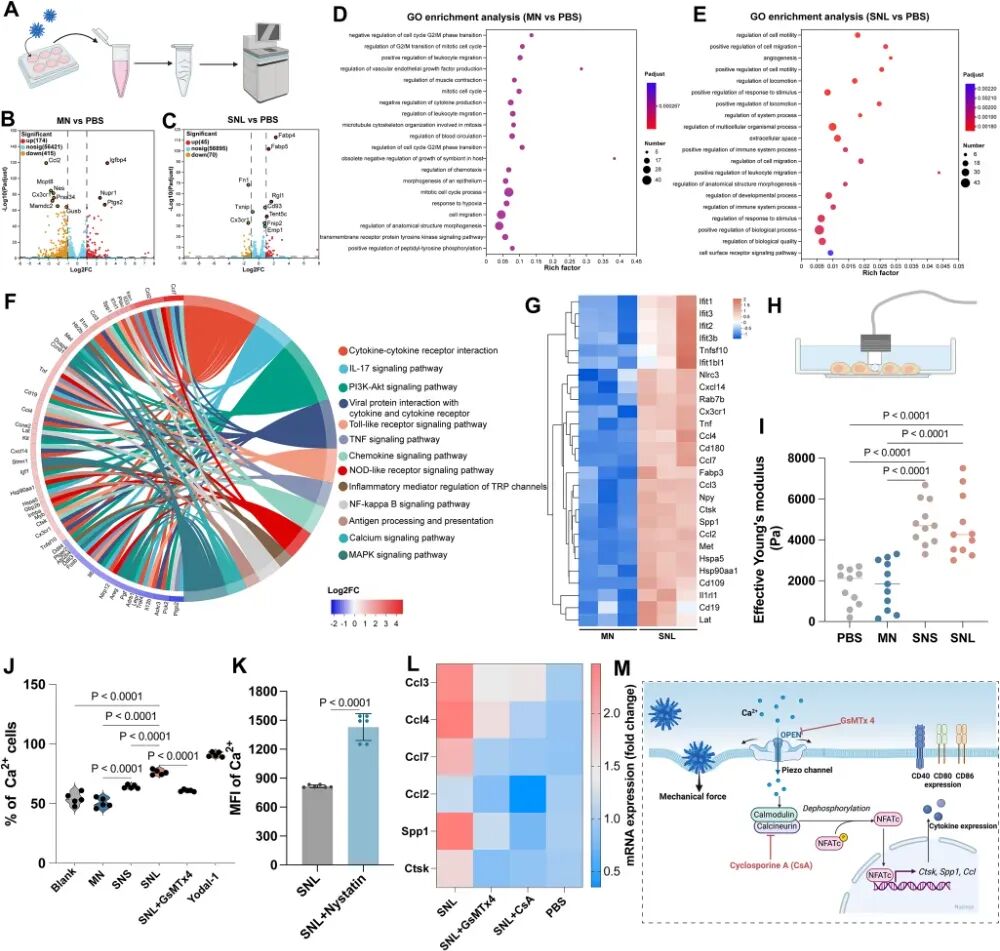
图2. 尖刺纳米颗粒通过机械刺激激活树突状细胞: (A) BMDCs分别经MN或SNL处理24 h后进行转录组测序的示意图。(B) MN vs PBS的差异表达基因火山图。(C) SNL vs PBS的差异表达基因火山图。(D-E) MN vs PBS(D)和SNL vs PBS(E)的GO富集气泡图。(F) SNL vs MN的差异表达基因KEGG富集弦图。(G) SNL vs MN上调基因的热图(Ctsk、Spp1、Ccl家族等)。(H-I) 纳米压痕测量细胞Young‘s模量示意图(H)及L929细胞经三种纳米颗粒处理1 h后的Young’s模量(I,n=11)。(J) 纳米颗粒处理后Piezo1激活诱导的钙内流检测(n=5)。(K) 抑制小窝蛋白(caveolin)后纳米颗粒刺激下的钙内流(n=6)。(L) 抑制Piezo1或钙调神经磷酸酶后SNL诱导的基因表达变化(n=4)。(M) SNL激活DC的机制示意图:机械激活Piezo1通道,促进钙内流,钙结合钙调蛋白激活钙调神经磷酸酶,使NFAT去磷酸化并入核,调控下游靶基因表达。
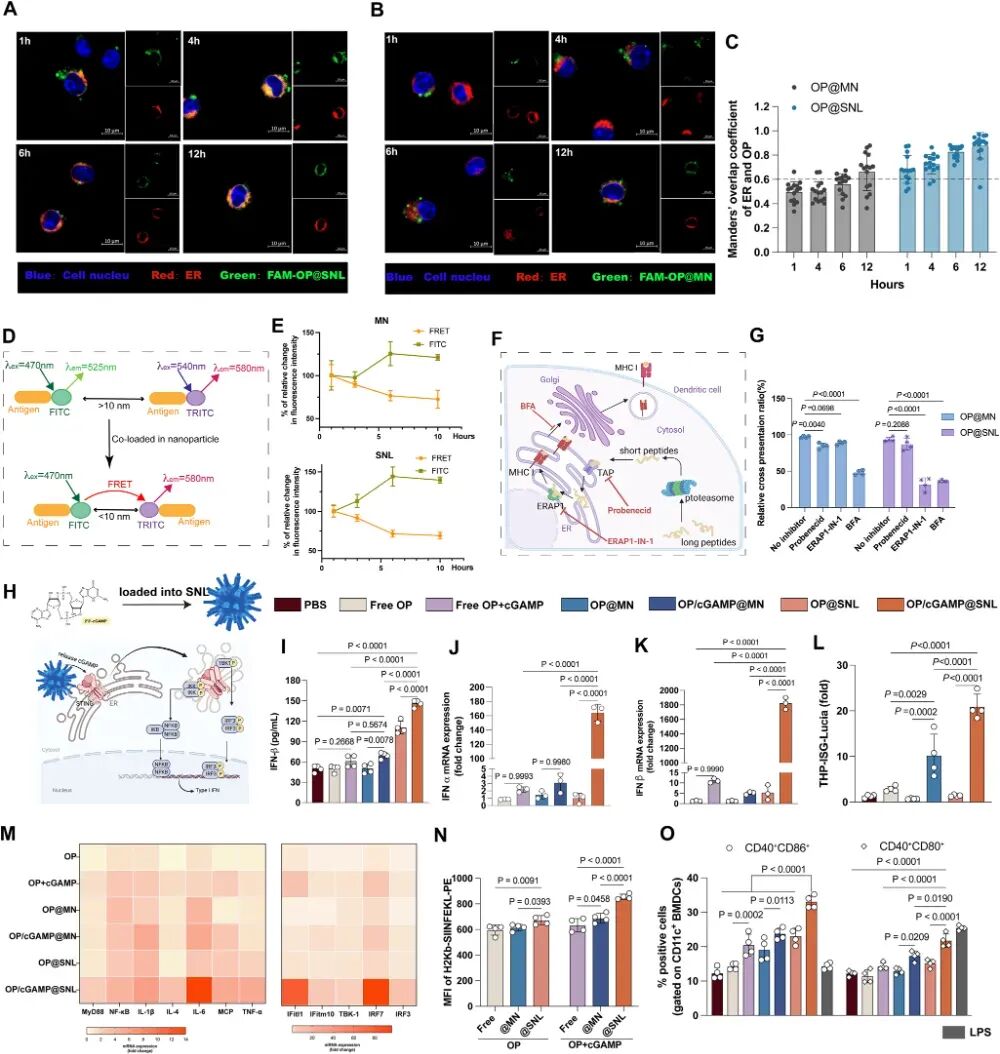
图3. SNL的细胞内运输行为及递送STING激动剂2‘3’-cGAMP的效率: (A-B) FAM-OP@MN(A)和FAM-OP@SNL(B)在DC2.4细胞中与内质网(ER)的共定位(绿色:FAM-OP,红色:ER,蓝色:细胞核)。(C) ER与纳米颗粒的Manders‘重叠系数定量(n=15)。(D-E) TRITC标记纳米颗粒负载FITC-抗原后,DC2.4细胞内FITC荧光强度及FRET效应随时间变化(n=4)。(F) APC加工和呈递长肽抗原的示意图(TAP、ERAP、ER-Golgi转运抑制剂)。(G) 抑制TAP、ERAP或ER-Golgi转运后,OP@MN或OP@SNL的相对交叉呈递比率(n=4)。(H) SNL负载cGAMP及STING信号通路示意图。(I-K) BMDCs处理后上清IFN-β分泌(I)及IFN-α(J)、IFN-β(K)mRNA表达(n=4或3)。(L) THP1-ISG-Lucia细胞中IRF通路活化倍数(n=4)。(M) BMDCs上清中IL-1β、IL-6、TNF-α分泌水平。(N) 抗原交叉呈递效率。(O) BMDCs成熟标志物表达。
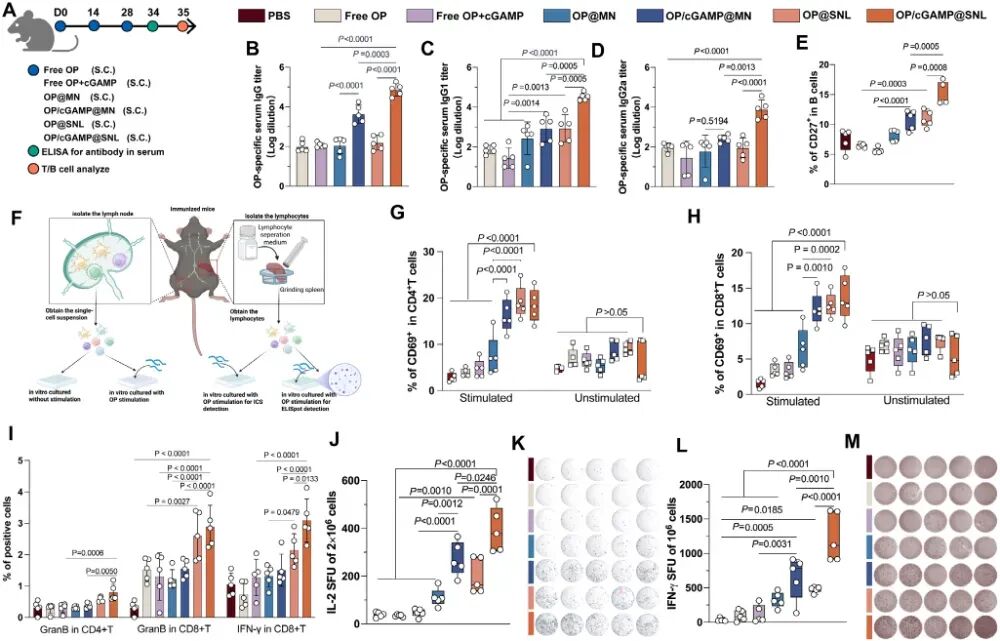
图4. SNL共载OP和cGAMP在体内触发免疫应答: (A) C57BL/6小鼠免疫方案示意图。(B-D) 第34天血清中OP特异性IgG、IgG1、IgG2a抗体滴度(n=5)。(E) 淋巴结中记忆B细胞比例(n=5)。(F) 抗原特异性T细胞活化检测示意图。(G-H) 体外有或无OP刺激下,淋巴结中CD4⁺ T细胞(G)和CD8⁺ T细胞(H)的特异性活化(n=5)。(I) 脾脏CD4⁺ T细胞和CD8⁺ T细胞胞内细胞因子染色(ICS)结果(n=5)。(J-M) 脾细胞经OP刺激后,ELISpot检测IL-2(J-K)和IFN-γ(L-M)分泌(n=5)。
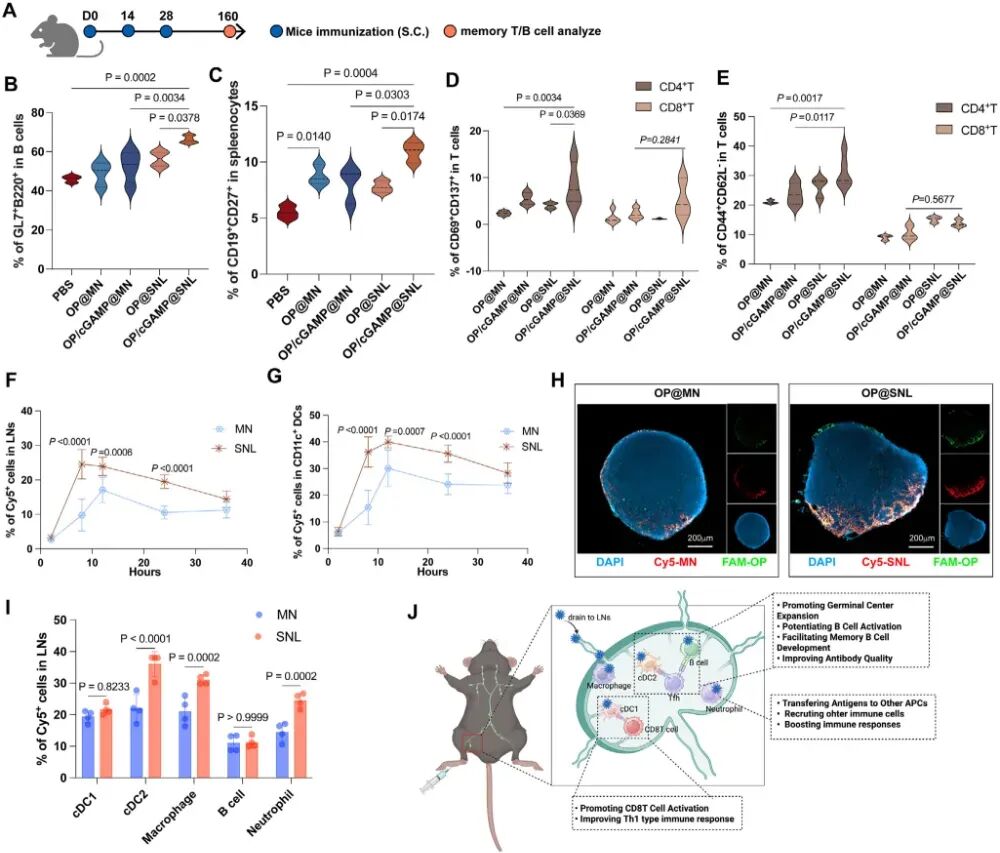
图5. SNL共载OP和cGAMP诱导长期记忆免疫应答: (A-E) 免疫后4个月检测记忆应答:生发中心B细胞(B)、记忆B细胞(C)、T细胞活化(D)和记忆T细胞(E)(n=3)。(F-G) 皮下注射后不同时间点,Cy5标记的MN和SNL在引流淋巴结中的积累效率(F)及在DC内的积累(G)(n=6)。(H) 注射12 h后淋巴结冰冻切片共聚焦图像,显示FAM-OP和Cy5-纳米颗粒的共定位(比例尺200 μm)。(I) 注射12 h后淋巴结内各细胞类型对MN和SNL的摄取情况(n=4)。(J) SNL皮下注射后引流至淋巴结并触发免疫应答的示意图。
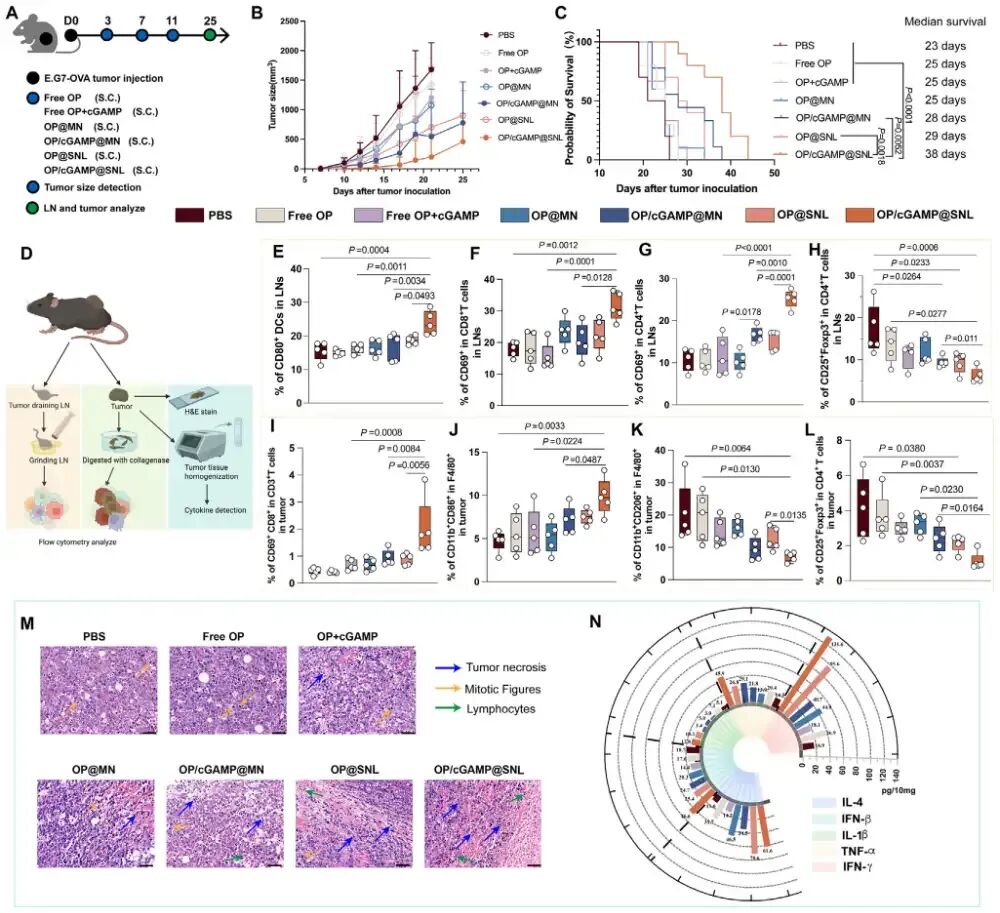
图6. SNL共载OP和cGAMP在EG7-OVA肿瘤模型中的抗肿瘤效应: (A) 治疗性疫苗接种方案示意图。(B) 平均肿瘤体积曲线(n=9)。(C) 生存曲线。(D) 第25天肿瘤引流淋巴结免疫应答检测示意图。(E-H) 肿瘤引流淋巴结中成熟DC(E)、活化CD8⁺ T细胞(F)、活化CD4⁺ T细胞(G)和Treg细胞(H)比例(n=5)。(I-L) 肿瘤微环境中活化CD8⁺ T细胞(I)、M1巨噬细胞(J)、M2巨噬细胞(K)和Treg细胞(L)比例(n=5)。(M) 肿瘤组织H&E染色显示坏死和淋巴细胞浸润。(N) 肿瘤组织内细胞因子水平(n=5)。
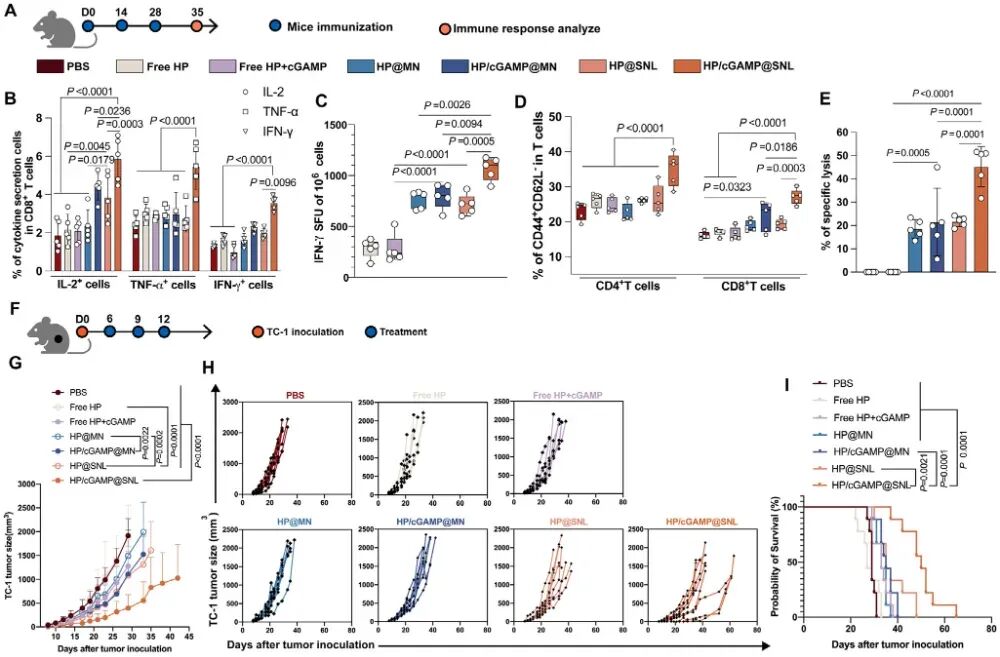
图7. SNL共递送HPV E7肽和cGAMP在宫颈癌模型中的效果: (A) 免疫方案示意图。(B) 脾细胞经HP再刺激后,CD8⁺ T细胞分泌TNF-α、IL-2和IFN-γ的水平(n=5)。(C) IFN-γ ELISpot结果。(D) 记忆CD4⁺ T细胞和CD8⁺ T细胞比例(n=5)。(E) 体内CTL杀伤效率。(F-I) TC-1肿瘤治疗模型:接种方案(F)、平均肿瘤体积(G)、个体肿瘤体积变化(H)和生存曲线(I)(n=9)。
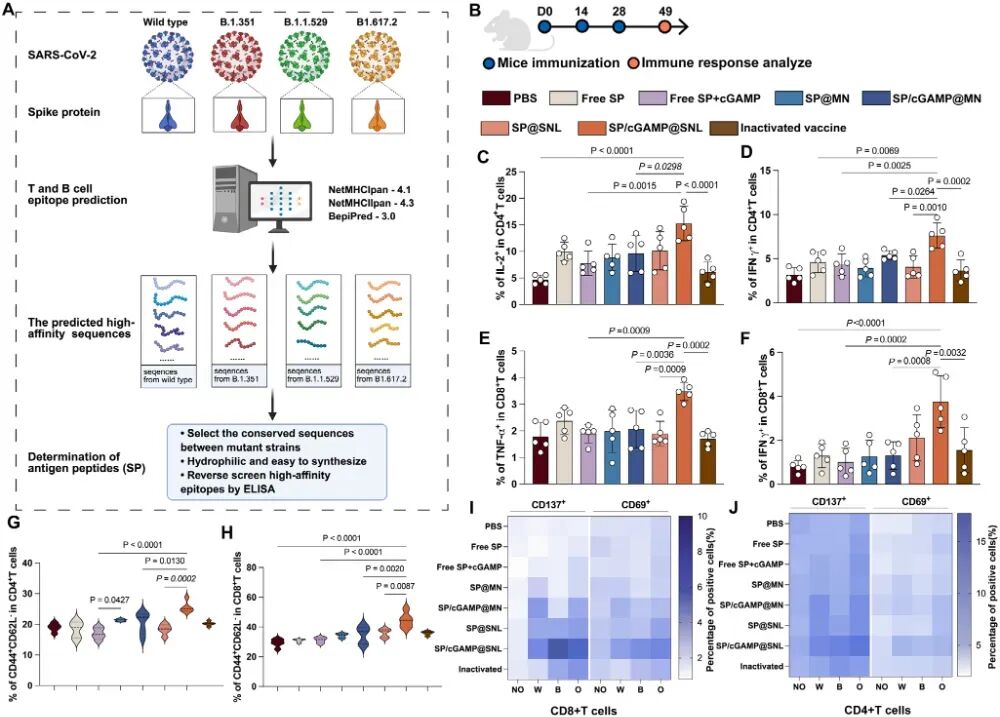
图8. SNL递送SARS-CoV-2相关肽在新冠模型中的交叉保护效果: (A) SARS-CoV-2抗原肽筛选流程示意图。(B) BALB/c小鼠免疫方案。(C-F) CD4⁺ T细胞分泌IL-2和IFN-γ(C-D),CD8⁺ T细胞分泌TNF-α和IFN-γ(E-F)水平(n=5)。(G-H) 再刺激后记忆CD4⁺ T细胞(G)和记忆CD8⁺ T细胞(H)比例(n=5)。(I-J) 用野生型(W)、Beta变异株(B)或Omicron变异株(O)刺突蛋白再刺激后,CD4⁺ T细胞(I)和CD8⁺ T细胞(J)的活化水平(n=4)。
结论
本研究首次证明了纳米颗粒表面的长尖刺形貌能够通过机械激活Piezo1通道,诱导钙内流,从而同时增强树突状细胞的活化和抗原向内质网的靶向递送。基于这一发现,团队发展了“淋巴结-APC-内质网”(LAE)级联递送策略,将抗原肽和STING激动剂cGAMP共包载于长刺介孔二氧化硅纳米颗粒(SNL)中。SNL不仅提高了淋巴结引流和APC摄取,还通过机械信号转导改变了囊泡运输路径,使抗原和佐剂共同富集于内质网,极大地增强了MHC I类交叉呈递和STING通路激活。在淋巴瘤和宫颈癌治疗模型中,SNL疫苗诱导了强效的CD8⁺ T细胞应答,延缓肿瘤进展并延长生存期;在SARS-CoV-2疫苗模型中,SNL疫苗激发了针对Beta和Omicron变异株的交叉保护性T细胞免疫。该工作揭示了纳米颗粒形貌作为疫苗设计关键参数的重要性,为开发下一代亚单位疫苗提供了通用平台。
原文链接:
https://doi.org/10.1002/adma.202573396