首页 > 医疗资讯/ 正文
线粒体是细胞内活性氧(ROS)的主要来源,在各类癌症中,ROS对细胞死亡途径(如焦亡)的调控起着至关重要的作用。PINK1介导的线粒体自噬能够有效清除受损线粒体并降低有害ROS水平,从而促进细胞存活。然而,PINK1和ROS在神经母细胞瘤中对焦亡的调控机制尚不清楚。
2025年3月31日,湖南大学王立明,中山大学卢盈颖和中南大学许浩东共同通讯在Autophagy在线发表题为“Inhibition of PINK1 senses ROS signaling to facilitate neuroblastoma cell pyroptosis”的研究论文。该研究表明,在神经母细胞瘤细胞中,抑制或缺失PINK1可敏化ROS信号,并通过BAX-caspase-GSDME信号通路促进焦亡。
具体而言,通过AC220抑制PINK1或敲除PINK1可损害线粒体自噬并增强ROS的产生,进而导致TOMM20的氧化和寡聚化,随后线粒体募集并激活BAX。激活的BAX促进CYCS(细胞色素c,体细胞)从线粒体释放到细胞质中,从而激活CASP3(caspase 3)。随后,激活的CASP3裂解并激活GSDME,诱导焦亡。此外,抑制或缺失PINK1可增强临床ROS诱导药物依他尼酸(EA)的抗肿瘤作用,从而在体内抑制神经母细胞瘤的进展。因此,本研究通过诱导焦亡为神经母细胞瘤提供了一种有前景的干预策略。

线粒体是众所周知的至关重要的细胞器,负责生成ATP和代谢物,以支持细胞活动。除了其在生物能量学和生物合成中的作用外,线粒体还在信号转导中发挥关键作用,包括调节细胞死亡、钙稳态以及活性氧(ROS)的产生和清除。作为信号细胞器,线粒体在内在凋亡的协调中扮演核心角色,这种凋亡可以由线粒体损伤诱导。此过程的关键事件是细胞色素c(CYCS)从线粒体释放到细胞质中。随后,释放的CYCS通过凋亡肽酶激活因子1(APAF1)促进半胱氨酸天冬氨酸蛋白酶9(CASP9)的激活;激活的CASP9进而裂解并激活CASP3,最终导致凋亡。此外,研究表明激活的CASP3可以裂解Gasdermin E(GSDME),将凋亡转化为焦亡[5,6]。GSDME由形成孔洞的N端结构域(GSDME-N)和自抑制的C端结构域(GSDME-C)组成。
在正常情况下,GSDME-C与GSDME-N结合,以维持GSDME的失活状态。然而,在应激条件下,激活的CASP3裂解GSDME,导致GSDME-N与GSDME-C解离。随后,释放的GSDME-N易位并在细胞膜上寡聚化,从而诱导焦亡。最近的研究报告指出,与胶质细胞相比,GSDME在神经元细胞(如SH-SY5Y)中高表达;此外,线粒体毒素可以激活CASP3依赖性的GSDME激活,并随后进一步损伤线粒体。尽管先前的研究已将GSDME与包括黑色素瘤和结直肠癌在内的几种癌症类型联系起来,但其在神经母细胞瘤及相关治疗中的作用仍不十分明确。最近的研究表明,高迁移率族蛋白B1(HMGB1)可能调节GSDME介导的焦亡,从而影响肿瘤进展和治疗反应。这凸显了进一步研究GSDME在神经母细胞瘤中作用的必要性。
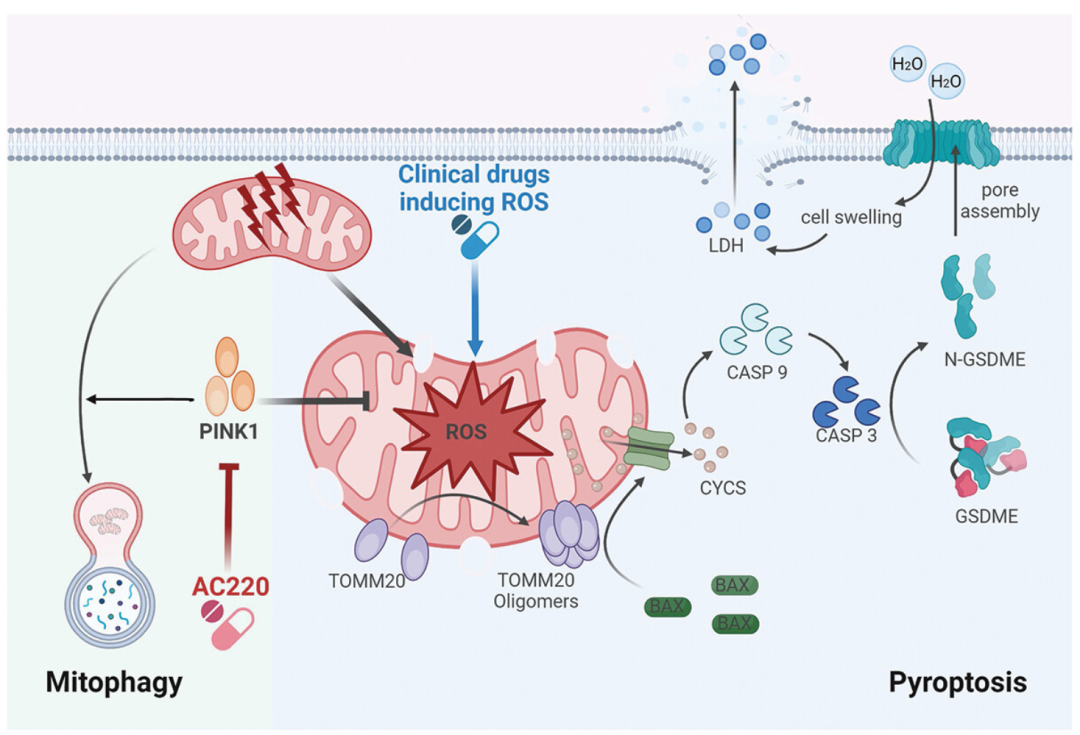
模式机理图(图片源自Autophagy)
ROS在癌细胞的存活和死亡中发挥重要作用。线粒体是氧化磷酸化过程中产生的细胞内ROS的主要来源,而线粒体ROS(mito-ROS)同时作为信号分子参与多种细胞过程,如衰老。在癌细胞中,mito-ROS作为第二信使参与致癌信号级联反应,促进肿瘤发展;然而,mito-ROS的过度积累可导致癌细胞DNA损伤和关键细胞通路中特定蛋白质的修饰,从而导致脂质过氧化、线粒体DNA(mtDNA)泄漏以及凋亡、坏死性凋亡和焦亡的诱导。在三阴性乳腺癌(TNBC)细胞中,四砷六氧化物通过抑制线粒体STAT3(信号转导子和转录激活子3)的磷酸化来增强mito-ROS水平,从而通过CASP3-GSDME途径诱导焦亡。在结直肠癌中,线粒体靶向的光动力治疗(PDT)刺激mito-ROS产生和p38磷酸化,导致通过CASP3介导的裂解激活GSDME,并随后诱导焦亡。因此,增强癌细胞中的mito-ROS作为一种治疗策略在临床癌症治疗中具有广阔前景。
线粒体自噬是选择性巨自噬/自噬的一种形式,它通过自噬-溶酶体途径专门消除受损或多余的线粒体。PINK1(PTEN诱导的激酶1)-PRKN介导的泛素驱动信号通路是线粒体自噬的关键机制。在健康线粒体中,PINK1通过其N端线粒体靶向序列(MTS)从细胞质转位到线粒体,并由TOMM和TIMM复合物导入线粒体。导入后,PINK1被线粒体蛋白酶裂解;随后,裂解的PINK1被逆向转位到细胞质并由蛋白酶体降解。相反,在线粒体损伤或去极化时,PINK1稳定存在于线粒体外膜(OMM)上,并在那里磷酸化泛素和PRKN[18,19]。激活的PRKN随后泛素化各种线粒体蛋白,招募自噬受体以随后将受损线粒体封装在自噬溶酶体内,随后与溶酶体融合并降解。据报道,线粒体自噬可以减弱mito-ROS的产生。
尽管线粒体自噬在神经退行性疾病(如帕金森病和阿尔茨海默病)中得到了广泛研究,但其在肿瘤发生和癌症治疗中的作用仍知之甚少。越来越多的证据表明,PINK1在癌症中具有双重作用。一方面,PINK1通过维持线粒体稳态和抑制功能障碍线粒体的积累来充当肿瘤抑制因子。胶质母细胞瘤、神经母细胞瘤和结直肠癌中已记录到PINK1的缺失,导致癌细胞存活和增殖增强。从机制上讲,PINK1介导的线粒体自噬负调节HIF1A/HIF-1α(缺氧诱导因子1亚基α)表达,从而抑制瓦伯格效应和炎症小体激活。此外,PINK1可以磷酸化TP53/p53(肿瘤蛋白p53),促进其核转位以抑制癌症干细胞(CSC)群。另一方面,PINK1可以通过增强CSC功能和维持肿瘤存活来促进癌症进展。具体而言,PINK1促进线粒体功能并激活MTOR(雷帕霉素激酶机械靶标)复合物2(MTORC2)-AKT信号通路,从而维持人脑肿瘤干细胞的功能完整性。此外,PINK1介导的线粒体自噬导致TP53降解,这反过来又进一步增强CSC活性和加速肿瘤进展。进一步研究PINK1的调节以及鉴定其激活剂或抑制剂可能为癌症治疗提供新的治疗见解。
神经母细胞瘤是儿童期最常见的颅外实体瘤,并且是儿科患者癌症相关死亡的主要原因。侵袭性神经母细胞瘤细胞对凋亡的抵抗,归因于CASP8(半胱氨酸天冬氨酸蛋白酶8)的缺失,这需要探索替代形式的程序性细胞死亡(如焦亡),以针对神经母细胞瘤制定有效的治疗策略。在本文中,作者通过证明PINK1的遗传和药理学抑制促进mito-ROS产生,从而激活BAX(BCL2相关X,凋亡调节剂)-半胱氨酸天冬氨酸蛋白酶-GSDME信号通路,并最终触发神经母细胞瘤细胞的焦亡性细胞死亡,来阐明神经母细胞瘤中焦亡诱导的机制。总体而言,作者的研究为治疗神经母细胞瘤提供了一种有前景的治疗方法。
原文链接:
https://doi.org/10.1080/15548627.2025.2487037
猜你喜欢
- 王小川:第一次做政协委员 建议让每个家庭都有家庭医生
- 咖啡+茶这么喝,健康益处能翻倍!柳叶刀子刊:经常喝咖啡,高风险人群患帕金森风险降8.6倍;搭配茶还有益大脑健康
- 年轻男子反复胸闷、夜里心悸醒来,以为是焦虑+胃食管反流!医生一做心电图:这是最容易“猝死”的遗传心脏病之一!
- 孕妇感染流感可以吃药吗?预防流感的好方法?
- J Ethnopharmacol:川续断皂苷D通过激活IGF1R/AMPK信号通路改善骨骼肌胰岛素抵抗
- 21-羟化酶缺陷反复IVF失败后成功妊娠1例
- 沙茶牛肉的做法-家常味炒菜谱
- Acta Haematol:随访期间与健康相关的并发症及其对血癌幸存者的影响——来自“血癌幸存者术后护理”(ABC)研究的结果
- Lancet Child Adolesc Health:抗精神病药物对儿童和青少年健康指标的影响
- 促进数据跨境有序流动
- 搜索
-
- 1000℃Nutrients:真实世界数据,纤维肌痛患者的饮食与运动自适应规律
- 1000℃D-二聚体升高诊治与管理专家共识(2026)
- 1000℃专家论坛|文良志:门静脉血栓的诊断和治疗
- 1000℃首例儿童NF2驱动型胸膜间皮瘤,多方法学检测锁定NF2双等位基因失活和14/22号染色体缺失,提示与成人胸膜间皮瘤不同
- 1000℃打破误区:干扰素追求CHB功能性治愈,HBsAg为何“不降反增”?
- 1000℃迷惑性极强的肝内病灶!影像表现疑点重重,最终病理竟查出两种不同肝脏恶性肿瘤
- 1000℃指南共识|原发性肝癌分子靶向药物相关蛋白尿中西医结合诊疗专家共识
- 1000℃Diabetologia:意大利北部社区 1~100 岁人群胰岛自身抗体与乳糜泻 TGA-IgA 的年龄分布及检测方法学验证
- 精J Child Psychol Psychiatry:12种罕见神经发育障碍儿童沟通能力谱系
- 精研究发现:爱吃辣的人,心血管病和癌症死亡风险都会显著降低
- 精Nursing in Critical Care:别再指责护士了!ICU 里被遗漏的护理,根源在系统而非个人
- 精Acta Obstet Gynecol Scand:罕见病女性的妊娠并发症与母婴结局,一项单中心434种罕见病的回顾性队列研究
- 精【爱儿小醉】儿科患者术前对流层臭氧暴露与围手术期呼吸系统不良事件之间的关系:一项单中心回顾性队列研究
- 精eBioMedicine:牙龈下微生物组与脑健康存在连续关联梯度,牙周炎或成认知衰退可干预靶点
- 精军事医学研究院《自然·通讯》:自适应IrPtCu纳米酶水凝胶实现耐药菌感染伤口序贯治疗
- 精能够逆转萎缩性胃炎的两个中成药,该怎么选择?
- 荐同时性多发性原发性肺癌,左右病灶分别为EGFR和ALK阳性,考虑淋巴结肿大仅局限左肺门及血浆EGFR阳性,采用奥希替尼联合化疗
- 荐40岁女性同时罹患卵巢支持细胞-间质细胞瘤和透明细胞乳头状肾肿瘤,WES等基因检测竟为阴性
- 荐椎管内麻醉使用止血药突发气道痉挛的抢救流程解析
- 荐女子肝区无任何不适,影像提示复杂囊性病变,层层鉴别后锁定罕见胆管源性囊性肿瘤
- 荐“绘”真报告 | 病理考虑为中枢神经细胞瘤,检出脑室外神经细胞瘤的特征性变异FGFR1-TACC1融合,辅助鉴别诊断
- 荐8岁女童出现男性化症状,竟是形似「性索-间质肿瘤」的卵巢「无性细胞瘤」所致,少见KRAS/CDK4共扩增或与侵袭性有关
- 荐17例病例分析揭示常见于中年人的色素性室管膜瘤临床特征与预后,分子检测可助力临床精准诊疗
- 荐Lancet Oncol:结直肠癌腹膜转移,围手术期化疗并非必选项
- 标签列表
-
- 星座 (702)
- 孩子 (526)
- 恋爱 (505)
- 婴儿车 (390)
- 宝宝 (328)
- 狮子座 (313)
- 金牛座 (313)
- 摩羯座 (302)
- 白羊座 (301)
- 天蝎座 (294)
- 巨蟹座 (289)
- 双子座 (289)
- 处女座 (285)
- 天秤座 (276)
- 双鱼座 (268)
- 婴儿 (265)
- 水瓶座 (260)
- 射手座 (239)
- 不完美妈妈 (173)
- 跳槽那些事儿 (168)
- baby (140)
- 女婴 (132)
- 生肖 (129)
- 女儿 (129)
- 民警 (127)
- 狮子 (105)
- NBA (101)
- 家长 (97)
- 怀孕 (95)
- 儿童 (93)
- 交警 (89)
- 孕妇 (77)
- 儿子 (75)
- Angelababy (74)
- 父母 (74)
- 幼儿园 (73)
- 医院 (69)
- 童车 (66)
- 女子 (60)
- 郑州 (58)