首页 > 医疗资讯/ 正文
解螺旋公众号·陪伴你科研的第3137天
助你科研思维再升级
Hippo通路目前已是生物研究领域尤其是肿瘤领域里家喻户晓级别的明星通路了,关于这条通路详细的机制和功能研究让很多科学家都成为了大佬,这也说明了这条通路的重要性。Hippo 信号转导是一条进化上非常保守的通路,可通过调控细胞增殖、凋亡及干细胞自我更新来控制器官大小。而Hippo 通路的失调会导致癌症发展。
2015年加州大学圣地亚哥分校药理系管坤良教授在Cell(IF= 66.85)杂志上发表了题为《Hippo Pathway in Organ Size Control, Tissue Homeostasis, and Cancer》的综述,系统性的介绍了Hippo通路的生物功能和作用,该文目前已被引用超过1600次。
管坤良教授本科毕业于杭州大学,博士学位取得于Purdue大学并在那里从事博士后研究。管坤良的研究工作主要在细胞信号转导、细胞生长、器官大小的控制和肿瘤发生。他在MAP kinase、mTOR和Hippo这几个领域都做出了开创性的工作,已经发表三百多篇文章,是分子生物学和遗传学领域里高引用研究者。

为了帮助大家更好的吸收综述的内容,本娃在阅读综述的时候,也给大家翻译了遍,希望能给大家带来有真正价值的“学术营养”。
摘要
20年来在多种模式生物中的研究明确了Hippo通路是器官大小和组织稳态的关键调节因子。通过抑制YAP和TAZ转录共激活因子,Hippo通路可调节细胞的增殖、凋亡和干性,以响应细胞外和细胞内的信号,包括细胞-细胞接触、细胞极性、机械信号、G蛋白偶联受体的配体和细胞能量状态。Hippo通路的失调对癌症发展有重要影响。对这一通路的功能和调节机制作进一步的研究将有助于揭开器官大小控制的奥秘,并鉴定癌症治疗的新靶点。
简介
多细胞生物体的出现是进化的里程碑。那些可确保组织和器官有适当的大小和形状并能满足其功能需要的机制是支持多细胞结构的最基本的机制之一。然而,尽管目前对“预设”器官大小背后的基本原理进行了深入研究,但我们对发育生物学中的这一基本问题还远没有一个清晰的认识。然而,对可控制器官大小的Hippo通路的研究则逐步揭示了这一谜团。
1995年,在果蝇中的两项研究发现,Warts(wts)基因的缺失会导致多种组织急剧过度生长。几年后,一系列研究表明,Salvador(sav)、Hippo(hpo)和作为抑癌因子的Mob(mats)突变型果蝇在促进组织过度生长方面所起的作用与wts突变体相似。Hpo、Sav、Wts和Mats在遗传和物理层面上相互作用,由于突变所引起的器官大小表型在其他已建立的发育通路中却没有发现,因此,它们被划分到一个新的信号模块——Hippo通路(由于hpo突变体有巨大的器官,故以此命名,类似于Hippo)。Yorkie(yki)是Hippo通路在器官大小调节中的关键功能效应因子,并很快也在与Wts相互作用的蛋白筛选中被发现。
Hippo通路在哺乳动物中高度保守。哺乳动物的Hpo、Sav、Wts、Mats和Yki的同源物分别是MST1/2 (也叫STK4/3)、Salvador(SAV1)、LATS1/2、MOB1a/b和YAP/TAZ (也叫WWTR1)(表一)。由于Hippo通路在调节器官大小方面的显著作用,以及它与组织再生和癌症的显著相关性,迅速引起了研究人员的广泛关注。在本文中,我们回顾了关于对Hippo通路如何组织、调节及其功能的研究,并讨论了一些重要的问题。
YAP/TAZ和Yki是Hippo信号传导的主要影响因子
在果蝇中,敲除Yki可以抑制hpo、sav或wts突变体的过度生长。在小鼠中,敲除Yap也能削弱因缺乏Mst1/2或其他上游调节因子而引起的过度生长。因此,Yki和YAP/TAZ是在进化上保守的Hippo通路关键效应因子。
Yki和YAP/TAZ也被认为可通过调控基因转录来介导Hippo通路的生物学功能。作为转录共激活因子,Yki和YAP/TAZ并不能直接与DNA结合,它们必须与DNA结合的转录因子相互作用才能调节靶基因的表达。在果蝇中,Scalloped (Sd)是YKI的转录因子和关键伴侣,它介导了Yki在组织生长中的功能。同样,在哺乳动物细胞中,TEAD家族的转录因子(TEAD1-4,Sd的同源物)是YAP的关键伙伴。研究证据表明,TEADs是YAP在转录调控过程中的主要伙伴。例如,有TEAD结合缺陷的YAP突变体会失去诱导YAP靶基因转录的能力,而嵌入了(knockin)TEAD的结合缺陷型YAP(Y94A突变体)则可模拟在皮肤和心脏中敲除YAP时的表型。此外,YAP和TEAD在基因组中的大部分位点都是共享的,当TEAD与VP16反式激活结构域融合时,所产生的基因表达谱与活性YAP所诱导的基因表达谱相似。TEAD1-4或Sd可以与类似于GTIIC序列(TGGAATGT或ACATTCCA)的共识序列结合,在GTIIC多联体(concatemers)控制下的转录报告因子现在被广泛用于测量Hippo通路的活性。除了TEAD1-4,YAP/TAZ也被证明可与其他转录因子相互作用,包括Smad、RUNX1/2、p63/p73和ErbB4,尽管这些转录因子在Hippo通路中的功能意义还不太明确。
在YAP/TAZ中存在一个较强的转录激活域,但在Yki中没有。然而,人类的YAP可以挽救果蝇Hippo通路过度激活导致的死亡,这也体现了其在功能上的保守性。对全基因组染色质结合状态评估的结果显示,除了占据靶基因的近端启动子外,YAP和TEAD主要通过与远端增强子相互作用来发挥其转录活性,这表明YAP(可能还有TAZ和Yki)可能会通过多种机制调节转录,包括招募通用(general)转录因子、修改表观遗传标记和调控染色质结构(图1)。事实上,研究证据表明,Yki可以与Brahma复合体、GAGA因子、核受体辅助因子6(NCoA6,Trithorax相关组蛋白甲基转移酶的一个亚单位)和中介复合体(Mediator complex)相互作用,而TAZ可以与染色质重塑复合体SWI/SNF相互作用。
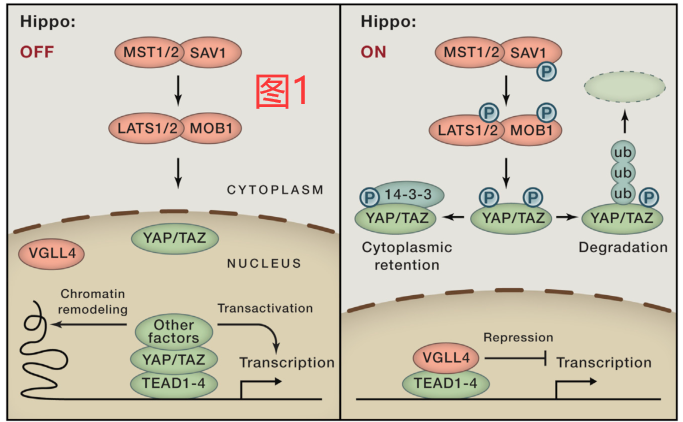
Yki和YAP/TAZ的转录活性在细胞核中受到Tgi (含有Tondu结构域的生长抑制因子)和VGLL4 (Tgi的同源物)的调节。Tgi可以直接与Yki竞争性结合Sd,导致Yki调节的转录受到抑制。当Hippo信号传导启动时,Tgi和Sd可形成复合体,导致转录抑制;相反,当Hippo信号传导关闭时,Yki可进入细胞核并从Sd中取代Tgi,从而促进Yki靶基因的表达和组织生长。在哺乳动物中,VGLL4同样可与YAP/TAZ竞争性结合TEAD (图1)。然而,YAP/TAZ是否仅通过解除VGLL4对TEAD的默认抑制而发挥作用则还没有得到证实。有趣的是,VGLL4的表达似乎可被YAP直接诱导miR-130a所抑制,而导致了YAP活性被放大。类似的机制也存在于果蝇中,其中Bantam(Yki诱导的miRNA)也可以抑制Tgi的表达。
尽管它们作为转录共激活因子的作用被广泛重视,但YAP/TAZ和Yki与特定因子结合时也可能会抑制某些基因的表达。例如,YAP/TAZ可以与NuRD复合物相互作用,导致转录抑制。此外,YAP也已被确认可通过调节miRNA加工酶Microprocessor或Dicer复合物而成为miRNA生物生成的调节因子,这表明YAP也具有非转录性依赖性的(transcription-independent)作用。因此,YAP/TAZ和Yki可能存在多种机制来调节基因的表达。
在果蝇和不同哺乳动物细胞类型中,有多项独立研究对YAP/TAZ和Yki超活化(或异位表达)下的基因表达特征进行了分析。然而,这些不同的基因分析研究之间的重合度并不高,这表明YAP/TAZ和Yki可能是以组织或细胞类型特异性的方式在调节靶基因表达。在果蝇中,Yki常见的靶基因包括Bantam、diap1和cyclin E,它们可能介导了Yki抑制细胞死亡和促进细胞增殖的作用。在哺乳动物中,CTGF是YAP激活的常用标志,许多参与细胞增殖、粘附和迁移的基因也被确定为YAP靶点,因此很可能是一组基因共同发挥了YAP/TAZ的生物学功能。
MST1/2和LAST1/2激酶抑制YAP的活性
在Hippo通路中,LATS1/2可直接磷酸化并抑制YAP/TAZ。有趣的是,YAP有五个LATS1/2靶向的共识motif(HXRXXS),其中四个motif在TAZ中是保守的。YAP在S127上的磷酸化可产生一个14-3-3的结合位点,并与胞质中的14-3-3螯合物YAP结合。此外,YAP在S381上的磷酸化可导致CK1δ/ε的磷酸化和磷酸二元体活化,促使SCFbeta-TRCP E3连接酶募集、泛素化和YAP的蛋白体降解。因此,通过调节YAP的亚细胞定位和蛋白质稳定性,LATS1/2具有对YAP活性在空间上和时间上的控制作用(图1)。LATS1/2对TAZ的调控方式类似,但可能由于TAZ的N端有一个额外的磷酸化降解子(phosphodegron,是一种磷酸化降解结构域),所以在TAZ的调控中降解有更重要的作用。Yki的亚细胞定位也受到Wts磷酸化和14-3-3结合的调节。然而,Yki中磷酸化降解子和磷酸化介导的降解机制并不保守。此外,YAP和Yki也被证明可通过自溶酶体通路降解,意味着YAP和Yki在胞膜动力学和相关细胞过程如自噬中具有潜在的作用。
LATS1/2可通过多种机制被MST1/2激活。MST1/2可在C端疏水基团上磷酸化LATS1/2,从而促进LATS1/2在其活性环中发生自磷酸化。此外,MST1/2对MOB1的磷酸化也可增强MOB1与LATS1/2自抑制结构域的相互作用,从而完全激活LATS1/2。此外,SAV1也可被MST1/2磷酸化,SAV1作为MST1/2的伙伴,在促进LATS1/2磷酸化方面发挥作用(图1)。在果蝇中,Wts受到Hpo、Sav和Mats的调节,其机制类似。
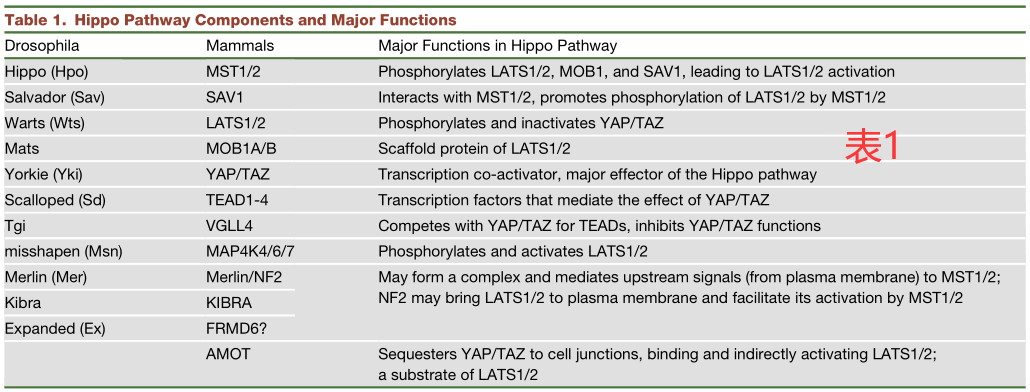
定位于细胞皮层(cell-cortex-localized)并与细胞骨架相互作用的三个蛋白,Merlin(Mer)、Expanded(Ex)和Kibra,它们可能是Hippo核心成分在顶端区域活性的支架,因为Sav、Hpo和Wts已被证明与Ex/Mer/Kibra之间存在物理性相互作用(表1)。此外,Ex/Mer/Kibra对Hippo通路的影响可能是由激酶Tao-1介导的,它可以直接磷酸化并激活Hpo。在哺乳动物细胞中,NF2 (Mer的同源物)似乎在调节LATS1/2的活性方面起着更直接的作用:它可以直接与LATS1/2相互作用,将LATS1/2招募到质膜上,并由MST1/2所激活(图2和3)。
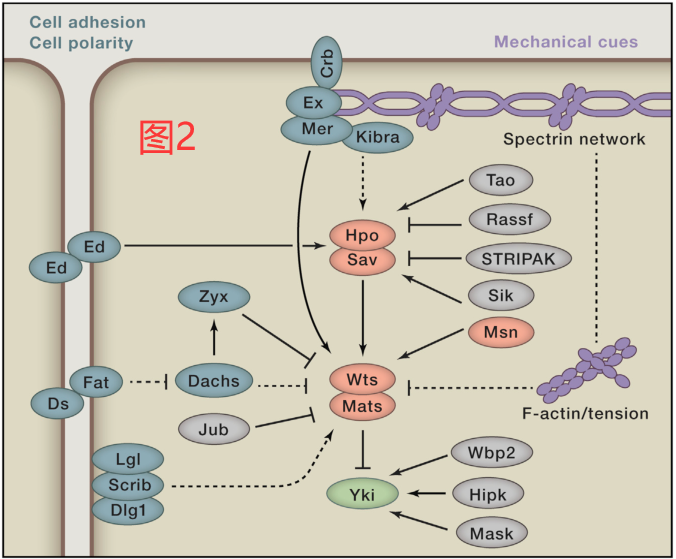
Hpo或MST1/2对Wts或LATS1/2的调节不是绝对必需的。在小鼠胚胎成纤维细胞(MEF)中,MST1/2的双重敲除并不能消除YAP的磷酸化,这表明还存在其他的Hippo样活性。事实上,最近在果蝇中的一项研究已经确定Misshapen(Msn)是另一个可激活Wts的激酶。这一机制在哺乳动物中也是保守的,因为MAP4K4(Msn的同源物)的过表达可促进LATS1/2的磷酸化,而MAP4K4的敲除则可诱导YAP报告因子的活性。除了MAP4K4,最近的两项研究都表明,许多MAP4K家族的激酶,包括MAP4K1/2/3/5和MAP4K4/6/7(果蝇的Msn),都可直接磷酸化并激活LATS1/2。这些激酶与MST1/2一起,可能以组织和信号特异性的方式在调节LATS1/2的活性。另外,其他的激酶,特别是一些STE20家族的成员,也有可能在不同上游信号或不同组织背景下激活LATS1/2(图2和图3)。
YAP/TAZ也被证明可被许多其他激酶磷酸化,如CDK1、JNK、HIPK、ABL和Src家族酪氨酸激酶,这也表明YAP/TAZ可被非Hippo通路激酶的机制所调节。
细胞极性和细胞粘附调节Hippo信号通路
在寻找Hippo通路的上游调节因子时,发现了许多参与细胞极性和细胞粘附的蛋白质。Echinoid(Ed)是果蝇的一种细胞粘附分子,它能与Sav相互作用并稳定Sav,从而激活Hpo。在哺乳动物细胞中,粘附和紧密连接处的几种蛋白,如AMOT、PTPN14和α-catenin,可以将YAP/TAZ截留(sequester)在细胞连接处。因此,细胞粘附和细胞间连接的形成可抑制YAP/TAZ的转录活性(图2和3)。
Crumbs(Crb)是顶端-基底极性的组成部分,可与Ex相互作用,这对Ex/Mer/Kibra复合物的顶端定位至关重要。此外,SCRIB可与MST1/2和LATS1/2相互作用,以促进激活LATS1/2。Fat作为在平面细胞极性中起关键作用的原钙粘蛋白,也可能通过调节Ex蛋白的水平和定位来激活Hippo通路 (图2)。然而,哺乳动物的Fat同源物似乎不是Hippo通路的主要调节因子。值得注意的是,细胞极性和Hippo通路之间可能存在间接联系,一些蛋白质,如Fat,也可能通过不同的机制调节细胞极性和Hippo信号。
细胞接触和机械信号调控Hippo信号传递
实体组织中的细胞可与邻近细胞及其细胞外基质(ECM)进行通信,并感知来自局部环境的持续物理信号。细胞-细胞接触被发现是调节Hippo通路的第一个信号。在稀疏培养(sparse culture)细胞时,YAP/TAZ主要定位在细胞核中,以促进靶基因的转录和细胞增殖。相反,在高细胞密度下,YAP/TAZ主要在细胞质中,起抑制生长的作用。众所周知,细胞在与周围的细胞发生物理接触后就会停止增殖,而细胞接触抑制的丧失是致癌转化的标志。因此,细胞密度对YAP/TAZ的调节提示Hippo通路在接触抑制、组织生长和肿瘤发生中起着重要的作用。
机械信号,如ECM硬度和细胞几何形状,也是YAP/TAZ的有效调节因子(图3)。当细胞生长在坚硬的基质上或散布在一个大的表面介质上时,YAP/TAZ会被激活。相反,当细胞定植在软基质上或被压缩到一个小区域时,YAP/TAZ被灭活。ECM的硬度和细胞的几何形状对细胞的增殖和分化很重要,而YAP/TAZ的活性在这些细胞过程中起重要作用。细胞的几何形状也被认为是细胞密度调节Hippo通路的一种机制:在低密度下,细胞平展,导致YAP激活;而在高密度下,细胞会采用圆形和紧凑的几何形状,导致YAP失活。为了进一步支持YAP/TAZ在机械传感中的作用,机械应变(mechanical strain)和剪切应力已被证明可刺激YAP/TAZ,而YAP的激活是机械应变所诱导的细胞周期再入的必要条件。在青鳉鱼(medaka fish)模型中,YAP也介导三维体形对组织张力的反应性变化。
细胞附着在ECM上是维持细胞存活的重要机制,当细胞脱离ECM时通常会失巢凋亡。细胞及其ECM的附着状态也可以调节Hippo通路的活性。YAP在细胞附着过程中被激活,也会在细胞分离(detached)时失活。组成性活化YAP的表达能促进分离细胞的存活,这提示有高YAP活性的癌细胞可能会避免失巢凋亡并发生转移。研究发现,在转移性前列腺肿瘤中,LATS1/2的表达会选择性地降低,而在原发性前列腺肿瘤中却没有这一现象。
可溶性因子调控Hippo信号
YAP/TAZ的主要功能是促进生长,许多促有丝分裂的激素和生长因子可通过G蛋白偶联受体(GPCR)作用以诱导细胞增殖(图3)。配体通过Gα12/13或Gαq/11偶联的GPCR发出信号,如溶血磷脂酸、凝血酶、血管紧张素II和雌激素,以激活YAP/TAZ;相反,配体也可通过Gαs偶联的GPCR和蛋白激酶A(PKA)发出信号,如肾上腺素和胰岛素,以抑制YAP/TAZ的活性。有趣的是,Gαq/11所激活的蛋白激酶C(PKC)既可激活又可抑制YAP,PKC的经典功能是激活YAP,而这种新的PKC则可抑制YAP。PKC调节YAP的功能差异在机制上解释某些细胞类型对PKC激活的特异性反应。GPCR是最大的膜受体家族,可介导各种生理或病理反应。GPCR对Hippo的调节表明Hippo通路与很多上游信号和生物功能存在紧密联系。
Wnt/β-catenin通路是发育和致癌的一个重要信号级联。包括Axin、APC和GSK3在内的破坏复合物(destruction complex)可导致β-catenin不断降解。Wnt刺激可干扰破坏复合物,使得β-catenin逐步积累。最近的研究表明,YAP/TAZ也可被各种Wnt家族的配体所激活。YAP/TAZ作为破坏复合物的组成部分,并以类似于β-catenin的方式受Wnt调节。然而,最近的一项研究表明,Wnt可通过Frizzled(一种类似于GPCR的Wnt受体)、Gα12/13、Rho GTPases和LATS1/2来激活YAP/TAZ。此外,作为SAV1和LATS1的支架蛋白,Apc的缺失可导致YAP的激活和肿瘤的发生。当然,可能还需要更多的研究来验证Wnt对YAP/TAZ的激活机制。
在哺乳动物细胞和果蝇中,表皮生长因子(EGF)和胰岛素通过Ras-Raf-MAPK信号级联或磷脂酰肌醇依赖激酶(PDK1)的介导,也可以调节YAP/Yki的活性。PI3K激活后也可由GSK3的直接磷酸化介导,使TAZ保持稳定。然而,在其他几项研究中并没有观察到EGF和IGF对YAP的明显影响,在有PI3K或AKT抑制剂或PDK1失活的胚胎细胞中,YAP的活性似乎是正常的。这些差异可能是由于实验环境或细胞类型不同所致,应通过进一步研究来澄清。
细胞代谢状态对Hippo通路的影响
最近有研究发现细胞代谢状态和Hippo通路之间存在联系(图3)。在能量剥夺的情况下,如葡萄糖饥饿,AMPK激活的蛋白激酶(AMPK)可以直接磷酸化YAP的S61和S94。YAP S94的磷酸化会消除YAP和TEADs之间的相互作用,从而抑制YAP的活性。此外,能量应激(energy stress)也可通过AMPK依赖性或非依赖性的方式上调LATS1/2的激酶活性以抑制YAP。AMPK可以磷酸化AMOTL1,这反过来又可促进LATS1/2对YAP的磷酸化。类似的机制也存在于果蝇中,其中Ampk使Yki失活,影响幼虫中央大脑和中央神经索的细胞增殖。在能源应激过程中,AMPK和LATS1/2都被释放出来以限制YAP的活动。这些发现还提示Hippo通路可能介导了二甲双胍的抗癌作用,目前已发现二甲双胍可激活AMPK。除AMPK外,葡萄糖也可能通过磷酸果糖激酶刺激YAP/TAZ和TEADs的相互作用,从而促进YAP/TAZ的活性。
YAP/TAZ的活性也与氧气的供应有关。在缺氧条件下,HIF1会刺激E3泛素连接酶SIAH1/2的表达。然后SIAH1/2促进LATS2的泛素化和降解,导致激活YAP/TAZ(图3)。此外,HIF1还可直接诱导TAZ的转录,而YAP与HIF1相互作用可稳定HIF1以增强HIF1靶基因的转录。
YAP/TAZ是细胞生长和增殖的有效刺激因子,而细胞的生长和增殖是个能量消耗的过程。AMPK对Hippo信号的调节表明,代谢状态可以作为YAP/TAZ促进生长活性的检查点。在营养匮乏或能量危机的条件下,YAP/TAZ的活性需要受到限制,以防止合成代谢过程引起的能量耗竭。氧气在细胞代谢中也起着关键作用,缺氧参与了多种病理过程,如癌症。缺氧和YAP/TAZ活性之间的联系表明YAP/TAZ在介导缺氧性的致癌作用中起一定的作用。
肌动蛋白细胞骨架整合上游信号
肌动蛋白细胞骨架和Rho GTP酶不仅在维持细胞形态方面很重要,在调节细胞增殖和分化方面也起着重要作用。调节肌动蛋白细胞骨架,如过量表达Rho GTPases或用C3毒素抑制Rho,可显著调节YAP/TAZ的活性。Rho GTPases和肌动蛋白细胞骨架动力学的变化作为一种机械信号与GPCR配体和细胞粘附作用都是调节Hippo通路的关键媒介(图3)。肌动蛋白细胞骨架的不同调节因子的缺失都会影响YAP和Yki的活性。例如,果蝇中肌动蛋白帽蛋白(actin-capping proteins)或Capulet基因(可抑制肌动蛋白聚合)的缺失都会导致Yki的激活和组织过度生长。同样,哺乳动物细胞中肌动蛋白帽蛋白或丝状肌动蛋白(F-actin)截断蛋白(cofilin或gelsolin)的敲除也可激活YAP。总的来说,Rho GTP酶的活性和F-actin似乎都能激活YAP/TAZ,而F-actin的不稳定则能抑制YAP/TAZ。
血影蛋白(Spectrin proteins)与短肌动蛋白丝联合可被组织成一个有弹性的多边形网状结构,排列在质膜的细胞内侧。在上皮细胞中,血影蛋白的网络定位极化,通常存在于顶端和基底层外侧。除了起支持细胞结构的作用外,血影蛋白网络还可传递来自细胞微环境中的多种信号以调节细胞功能。最近,有三项独立研究揭示了血影蛋白对Hippo通路的调节作用,因为在果蝇的多个组织中,血影蛋白网络的破坏导致了Yki的激活和组织生长。在哺乳动物细胞中也观察到了类似于血影蛋白调节YAP的作用(图2和3)。因此,血影蛋白或相关肌动蛋白丝可能是整合上游信号(如机械信号)的主要节点。
尽管肌动蛋白骨架的作用很重要,但目前仍不清楚肌动蛋白细胞骨架是如何调节Hippo通路核心激酶的活性。有一种可能是多个Hippo通路的组成成分通过肌动蛋白介导的机制在顶端区域富集以促进信号转导,而肌动蛋白重塑可能会加强或破坏Hippo通路成分的集聚。在另一种情况下,Hippo通路可能受到收缩性肌动蛋白和细胞张力的调节(图3)。抑制与张力有关的酶,如非肌动蛋白(Blebbistatin, ATPase抑制剂)、ROCK(Y27632,ROCK抑制剂)和肌动蛋白轻链激酶(ML-7,MLCK抑制剂),都会抑制YAP/TAZ。然而,Hippo通路激酶是如何感知张力的,目前还不完全了解。用小分子处理细胞以抑制细胞张力,也可能会影响肌动蛋白的动态,因此很难区分肌动蛋白重塑和肌动蛋白张力所产生的作用。然而,有人提出,在果蝇中,Ajuba(Jub)在张力诱导的Yki调节中起关键作用。在对高张力的反应中,Jub通过与α-catenin相互作用将Wts招募到细胞间的连接处,从而抑制Wts活性。此外,机械应变(mechanical strain)时JNK的激活也可抑制LATS1。
尽管最初有研究报道机械信号对YAP的调节独立于LATS1/2,但最近的研究表明LATS1/2也参与了肌动蛋白细胞骨架对YAP/TAZ的调节。这种差异可能是由于在早期研究中,LATS1/2并没有在敲除实验中被完全去除。那些支持LATS1/2起了作用的研究发现,在肌动蛋白细胞骨架重排时,LATS1/2的磷酸化状态和体外激酶活性会明显受到调节,其动力学与YAP/TAZ磷酸化的动力学相似。血影蛋白或细胞的附着/分离对Yki/YAP/TAZ的调节依赖于Wts和LATS1/2。此外,在小鼠胚胎发育的前期,LATS1/2对靶向肌动蛋白细胞骨架的小分子在调节YAP时至关重要。总之,机械信号很可能通过Wts和LATS1/2来影响Yki和YAP/TAZ的活性和功能。
负反馈调节和串联
Yki或YAP/TAZ的高活性,特别是长期性的高活性,可导致组织的过度生长或癌症。因此,必须严格控制Hippo信号的生理波动,以避免有害影响。在果蝇中,一个内置的负反馈环可以微调Hippo通路,其中Yki的激活可启动上游调节因子的表达,包括Four-jointed、Ex、Mer、Kibra和Wts。一致的是,在哺乳动物细胞中也存在类似的负反馈机制。YAP/TAZ可直接诱导NF2、LATS2、可能还有MST1的转录,导致激活LATS1/2和抑制YAP/TAZ。此外,YAP/TAZ还可诱导其负向调节因子AMOTL2的表达。这种负反馈回路对于维持YAP/TAZ在受到刺激时有适当瞬时活性至关重要。通过这些机制,YAP和TAZ相互拮抗,并可能为Hippo通路活性的波动提供缓冲,以确保组织中的动态平衡。当这种负反馈被破坏时,Hippo通路的失调就可能导致肿瘤的发生。
Hippo通路的调控和功能会因为Hippo通路可与许多其他发育信号通路串联而变得复杂。除了Wnt,YAP/TAZ也可能受到SHH信号(sonic hedgehog)的调节。另一方面,YAP/TAZ也可以调节Wnt、SHH、TGF-β、JAK-STAT、EGFR和Notch通路配体的表达。信号的广泛串联可能创造了一个富含各种因素的微环境,进而又可通过自分泌或旁分泌机制以细胞自主和非自主的方式调节细胞命运。
胚胎早期发育中的Hippo通路
YAP/TAZ在胚胎早期发育中至关重要。尽管敲除了TAZ基因的小鼠依然可以存活,但敲除了YAP的小鼠在E8.5时就会死亡,当YAP和TAZ都被敲除时,胚胎在桑椹胚(16-32个细胞)之前就会停止分裂。因此,YAP/TAZ在早期发育中的作用存在部分重叠。
胚胎发育过程中的第一次细胞命运分化(cell fate specification)发生在着床前阶段,胚胎形成滋养层(TE)和内细胞团(ICM)。滋养层由囊胚的外层细胞组成,并形成胚胎外组织,而内细胞团由囊胚的内层细胞组成,并产生胚胎本身和其他组织。TE和ICM的形成主要是由于桑椹胚细胞的位置或极性,其中的内部和非极性细胞形成ICM,而外部和极性细胞形成TE。早在16细胞阶段,YAP/TAZ就表现出了内外细胞之间不同的亚细胞定位,这种差异一直持续到囊胚阶段。YAP/TAZ在TE和ICM中的不同分布导致形成了不同的基因表达特征(尤其是TE特异性诱导的基因,如外细胞中的Cdx2),从而导致了细胞命运的分化。事实上,敲除了TEAD4的小鼠胚胎不能发育出TE细胞,所有细胞都分化为ICM。另一方面,剥夺LATS1/2、NF2或AMOT/AMOTL2都会使所有细胞变成TE系细胞,这些胚胎都未能发育出ICM衍生组织。以上结果表明,Hippo通路在早期胚胎细胞分化中起着关键性的作用。
Hippo通路在器官大小控制和组织稳态中的作用
Hippo通路最知名的生理功能就是调控器官大小。在果蝇中,Hippo通路的激酶(hpo和wts)或上游调节因子(ex、mer、kibra、ft等)的突变可导致器官如眼睛、翅膀或其他附属物过度生长,yki的转基因表达(transgenic expression)也会产生类似的表型。组织/器官的增大主要是由于Yki诱导的细胞增殖和存活。
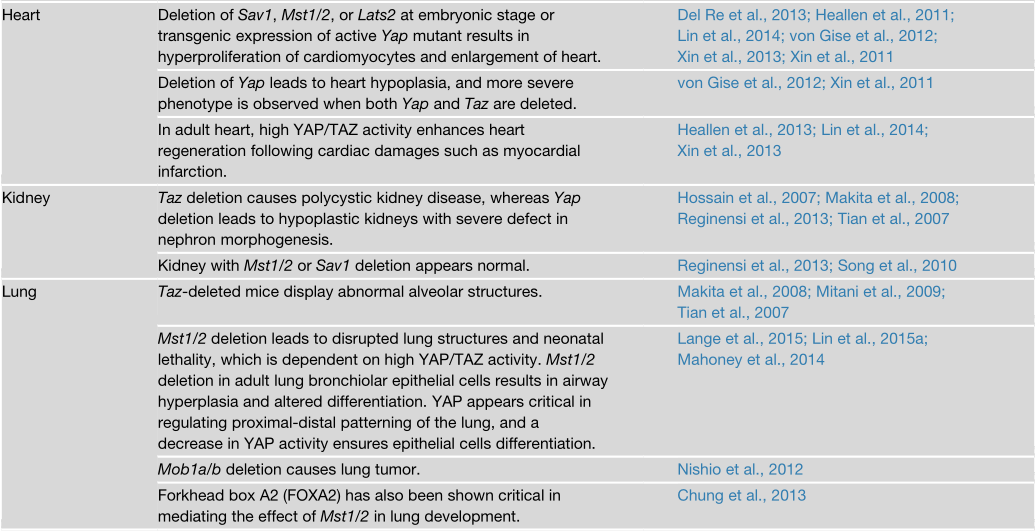
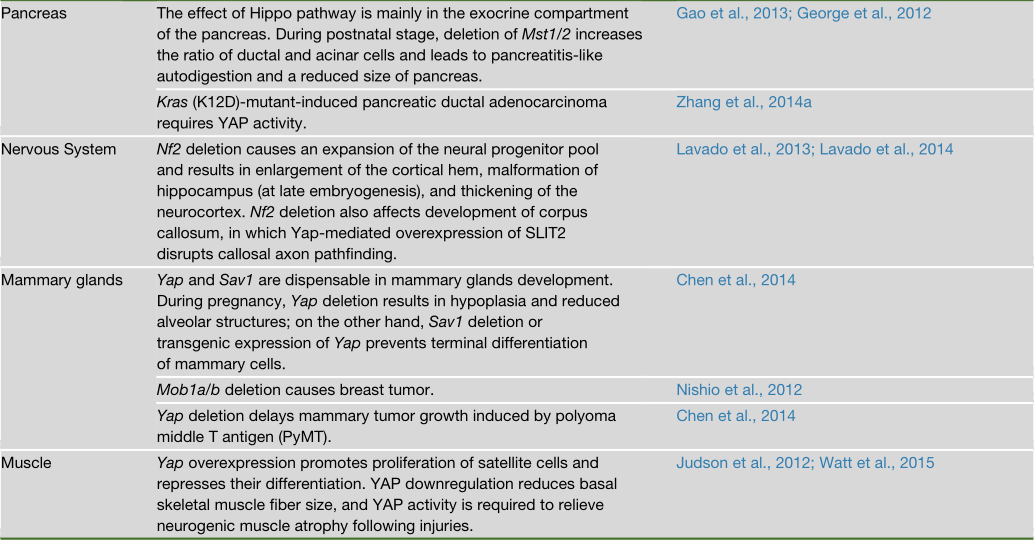
Hippo通路对器官大小的影响在哺乳动物中高度保守,在小鼠中进行的多项研究都表明了这一点(表2)。例如,小鼠肝脏特异性的Yap转基因表达会导致肝脏急剧增大。一旦Yap的过表达被关闭,肝脏就会通过细胞凋亡恢复到正常大小。同样,在肝脏中特异性地敲除Mst1/2、Sav1或Nf2也会导致肝脏增大。当Sav1、Mst1/2或Lats1/2被敲除时,小鼠的胚胎心脏也会增大,而且心肌细胞的增殖或凋亡对Yap的遗传操作很敏感。
然而,并非所有的器官都对Hippo通路的突变同样敏感。例如,敲除Mst1/2可导致肝脏、心脏、胃和脾脏急剧过度生长,但肾脏、肺和四肢却没有。其中一个可能的解释是,在这些组织中存在不依赖于MST1/2的YAP/TAZ调节因子。在乳腺和肠道中,Yap的组织特异性缺失不会导致组织结构或大小有任何缺陷。在小鼠肝脏中敲除Yap基因会导致胆管缺陷,但基本没有导致肝脏变小。这种效应的缺乏可能是由于TAZ的存在,TAZ应该是在YAP缺失时由于丧失反馈抑制而被激活的。另外,可能在某些器官中,Hippo通路或YAP/TAZ活性对器官大小的控制作用可以忽略不计。然而,Hippo通路对大小控制的器官特异性影响可能表明,不同器官的大小调节机制是因为利用了不同的调控原则。例如,器官的大小可以由分化细胞的增殖或预先分配的祖细胞数量来决定。
在器官大小决定过程中,Hippo通路上游的关键生理信号还没有被确定。机械力或张力可能会因为器官的生长而改变,并可能在器官生长到最终大小时抑制YAP/TAZ。另外,器官的大小可能是由可溶性因子通过自分泌/旁分泌机制所限制/诱导的,而这种因子的浓度是由器官大小所控制。但这两种模式并不互斥,需要进一步研究以理清Hippo通路是如何感知生理信号以调节器官大小的。
研究人员在多种组织的干细胞或祖细胞中发现YAP/TAZ的活性高,表明YAP/TAZ在干细胞的维持中起作用。例如,YAP在基底祖细胞和位于隐窝基底的肠道干细胞中具有高度的核性(highly nuclear)。通过转基因表达YAP或敲除上游调节因子来激活YAP,通常会导致祖细胞的增殖,细胞分化抑制,以及靶标组织如肠道、肝脏、皮肤和神经系统的过度增生。
YAP/TAZ对细胞增殖和干细胞扩增的作用表明YAP在正常组织发育和稳态中具有关键功能。事实上,Yap的组织特异性缺失会导致心脏、皮肤和肾脏的异常。然而,乳腺和肠道在Yap缺失后仍然相对正常。这些发现表明,YAP是小鼠某些(而不是所有)组织的发育和稳态所必需的。在人类中,TEAD1突变见于Sveinsson脉络膜视网膜萎缩症(一种以脉络膜视网膜变性为特征的疾病)和Aicardi综合症(一种先天性神经发育障碍)中。此外,YAP的功能缺失突变在孤立性和综合征性的视裂隙闭合缺陷中均有发现。因此,TEAD介导的YAP转录活性缺失会在人类的一些变性相关疾病中起作用。

尽管一些组织的发育和正常平衡并不需要YAP,但YAP的活性对某些类型组织的损伤后再生至关重要。例如,尽管乳腺发育正常,但Yap的缺失还是会严重影响怀孕诱导的乳腺组织生长。同样,在野生型小鼠中,用葡聚糖硫酸钠(DSS)诱导结肠炎后,肠道仍可以有效地再生;然而,在条件性敲除了Yap的小鼠中,再生能力受到严重阻碍。类似的结果也在果蝇中肠的DSS诱导损伤后再生中得到了验证。正常情况下,肝脏受损后会有效再生。例如,在切除部分肝脏后,肝细胞开始增殖并在几天内恢复肝脏的质量,在这个过程中,YAP活性会被诱导,这很可能是完整的肝脏再生所需要的。与肠道和肝脏相比,成人心脏的组织再生是非常有限的。然而,Hippo通路的失活或Yap的转基因表达可以恢复一些心肌的再生能力,尽管其效率很低。相反,心脏特异性缺失Yap会阻碍新生儿心脏的再生。总之,这些结果表明,YAP在多种组织的再生过程中起重要作用。
癌症中的Hippo信号通路
YAP的长期激活,如在小鼠肝脏中转基因表达Yap,可导致细胞转化和肿瘤的发展,这体现了Hippo通路在癌症发生和发展过程中的作用。表2总结了小鼠模型中Hippo通路在肿瘤发生过程中起作用的研究证据,总体上支持YAP/TAZ的致癌作用,以及Hippo通路上游成分的抑癌功能。
YAP的促肿瘤活性在很大程度上依赖于TEAD介导的转录过程,因为YAP诱导的肝癌可被显著负性的(dominant-negative)TEAD(能够隔离YAP和TAZ)的表达完全抑制。在细胞水平上,YAP的激活对细胞增殖、生存、迁移和侵袭都非常重要。YAP或TAZ的高活性使细胞能够避免接触抑制和失巢凋亡,并支持非停泊性生长(anchorage-independent growth,不依赖于支持物的存在)。YAP可诱导表达ZEB1/2,刺激上皮向间质转化(EMT),这是肿瘤转移的一个关键步骤。此外,YAP还能够促进基因组的不稳定性,而TAZ还是维持乳腺癌干细胞的自我更新和肿瘤诱导能力所必需的。
越来越多的证据表明,在许多人类癌症中Hippo通路会发生失调。在许多癌症类型中都有观察到YAP/TAZ的表达升高或核富集,包括肝脏、乳腺、肺、结肠、卵巢等等。然而,大多数具有高YAP/TAZ活性的癌症与Hippo通路的基因突变无关,而且人类癌症中Hippo通路组分的总体基因突变率都相对较低(表3)。
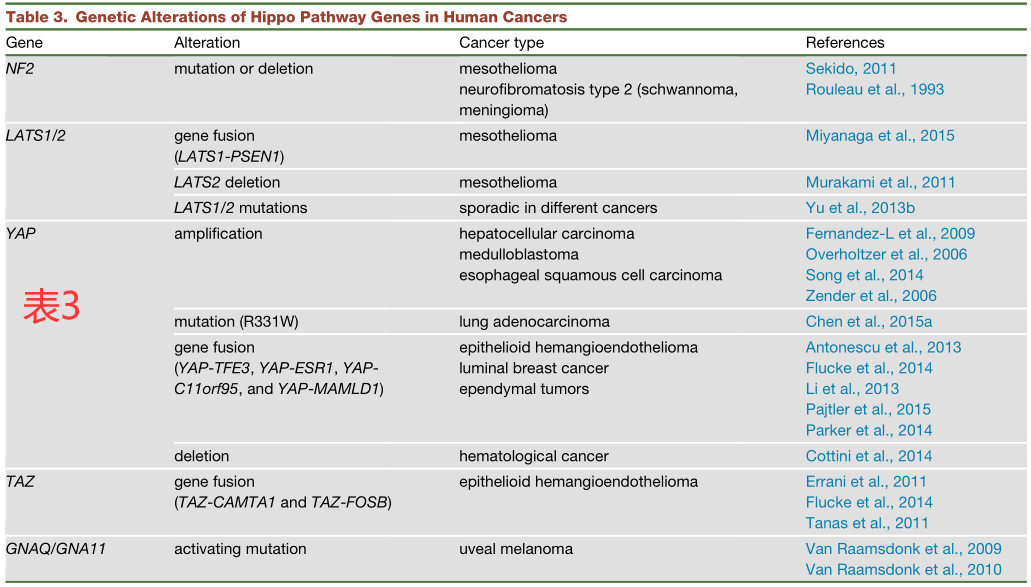
在与癌症相关的Hippo通路突变中,NF2是一个很好的例子,它可导致神经纤维瘤2型病变,包括神经鞘瘤和脑膜瘤。此外,在40%-50%的恶性间皮瘤中也能观察到NF2的失活突变。另外,即使是Yap的杂合性缺失也能完全阻断敲除Nf2所诱导的小鼠肝脏肿瘤的发生,这表明激活YAP是介导Nf2突变致瘤潜力的主要机制。
YAP的基因扩增可能导致了一部分肝细胞癌、髓母细胞瘤和食道鳞状细胞癌。涉及YAP或TAZ的基因融合也已在人类癌症中被发现。几乎所有的上皮性血管内皮瘤中都含有TAZ-CAMTA1、TAZ-FOSB或YAP-TFE3的基因融合。此外,在一部分上皮瘤中也发现了YAP基因与MAMLD1或C11orf95的融合。值得注意的是,在上皮样血管内皮瘤和室管膜瘤中,所有的YAP/TAZ融合蛋白都保留其N端TEAD结合域,但失去了C端反式激活结构域。这些观察结果表明,YAP的融合基因仍然可能会结合并激活依赖于TEAD的转录程序,以促进肿瘤的发生。事实上,携带YAP-C11orf95融合基因的神经干细胞移植到小鼠体内后还可以有效地形成脑瘤。此外,据报道,家族性的YAP点突变(R331W)也与肺腺癌的高发生率相关。
不正常的GPCR信号传递也常常导致肿瘤的发生,因此GPCR失调后有可能通过激活YAP/TAZ而导致癌症。GNAQ-或GNA11-(分别编码Gαq或Gα11)的激活突变已在80%的葡萄膜黑色素瘤中有发现,并可作为驱动突变发挥作用。最近的研究表明,在有GNAQ或GNA11突变的葡萄膜黑色素瘤中,YAP会被构成性激活,而高YAP活性有助于肿瘤的生长。在小鼠中,皮肤干细胞中Gnas(编码GαS)的缺失会导致基底细胞癌变,而这一过程部分依赖于YAP。此外,一种病毒性GPCR的表达也可诱导卡波西肉瘤的肿瘤发生,其中YAP/TAZ也起到了关键作用。
在不同癌症中也有零星发现LATS1/2的突变或基因融合,这可能会导致YAP/TAZ的激活(表3)。此外,与其他癌症相关信号通路的串扰也可能导致Hippo通路成分没有突变的癌症仍具有高活性的YAP/TAZ。例如,KRAS、APC和LKB1的突变都已被报道可激活YAP/TAZ。
YAP/TAZ的活性也与肿瘤的耐药性和复发有关。具有高YAP/TAZ活性的乳腺癌细胞可对诸如紫杉醇、5-FU和多柔比星等药物表现出抗性。此外,具有高YAP活性的肺癌和结肠癌细胞对RAF和MEK的靶向治疗也具有抗性。他莫昔芬通常用于治疗雌激素受体(ER)阳性的乳腺癌;然而,有一些ER+的乳腺癌患者对他莫昔芬并不敏感。最近,研究发现他莫昔芬可通过刺激膜雌激素受体GPER(一种GPCR)来激活YAP/TAZ。因此,他莫昔芬或雌激素对GPER的激活作用可能有助于肿瘤的生长和耐药。YAP基因的扩增也与KRAS驱动的结肠癌和胰腺癌复发有关。因此,抑制YAP/TAZ不仅可以针对肿瘤的发生和发展过程,而且有可能使癌细胞对化疗敏感,防止癌症复发。
值得注意的是,与其在大多数实体瘤中的致癌功能相反,YAP在血液肿瘤中似乎起着抑癌的作用。YAP基因座在血液肿瘤中经常缺失,表达YAP或抑制MST1都会导致生长抑制和凋亡增加。目前,YAP在血液肿瘤中抑癌功能的基本机制还不是很清楚。
治疗性靶向Hippo信号通路
Hippo通路的核心是一个激酶级联反应,而蛋白激酶通常是可以成药的(druggable)。因此,可以开发MAP4K4、MST1/2或LATS1/2的抑制剂来诱导YAP/TAZ的活性,以促进伤口愈合、组织修复或再生等过程,并可能用于治疗退行性疾病(图4)。例如,暂时性的抑制MST1/2或LATS1/2可能会促进心肌再生或存活,从而使心脏病患者受益。MST1/2或LATS1/2抑制剂也有可能用于治疗血液肿瘤。
一般来说,MST1/2和LATS1/2是抑癌因子,抑制MST1/2或LATS1/2可能在大多数情况下都会促进肿瘤生长。因此,抑制YAP/TAZ的活性将提供一个新的、有吸引力的抗癌策略。YAP/TAZ的功能主要是由TEAD介导的,因此能够破坏YAP/TAZ-TEAD相互作用的小分子就可以作为YAP/TAZ抑制剂来发挥作用。事实上,卟啉家族的分子,特别是verteporfin,能够破坏YAP/TAZ和TEADs之间的相互作用,verteporfin还可以阻断YAP/TAZ靶基因的转录,抑制YAP过表达或NF2失活所诱导的小鼠肝脏过度生长。然而,verteporfin的普遍细胞毒性和低水溶性限制了它的前景。根据YAP-TEAD和VGLL4-TEAD复合物的结构信息,研究人员设计了一种被称为“‘super-TDU”的多肽来阻断YAP-TEAD的相互作用,并已被证明能可抑制小鼠模型中的肿瘤生长。

设计可直接激活蛋白激酶的因子是个很有挑战性的策略。然而,LATS1/2可能会被靶向其上游调节因子的分子间接性地激活。目前第一个被发现对YAP有抑制作用的小分子(多巴酚丁胺)是一种GPCR受体拮抗剂。此后,许多YAP/TAZ的间接抑制剂也被发现,包括磷酸二酯酶抑制剂rolipram和ibudilast。Rho家族的GTP酶对LATS1/2有很强的抑制作用,而膜定位对Rho细胞功能有重要影响。事实上,甲羟戊酸代谢通路抑制剂(他汀类药物)可以阻断Rho GTPases的膜转位并间接抑制YAP/TAZ的活性。检验这些药物是否能有效抑制小鼠模型中肿瘤的生长,并进行流行病学研究以确定使用他汀类药物或rolipram的患者是否癌症发病率较低,这将会是一项有趣的研究。鉴于Hippo通路在调节细胞增殖和组织稳态方面的重要功能,它将是一个令人兴奋同时也需要进一步探索的癌症治疗领域。
未解决的问题
尽管Hippo通路的研究进展迅速,但一些关键问题仍然没有答案,而且新的问题正在涌现。下面列出了Hippo领域的一些关键问题。
1.调控MST1/2活性的分子机制是什么?许多上游信号可以调节LATS1/2的磷酸化和激酶活性。然而,MST1/2的磷酸化及其激酶活性都没有受到上游信号的显著调控。果蝇的misshapen(哺乳动物中MAP4K的一个成员)在Wts的上游起作用。另外,MST1/2和MAP4Ks是否会介导不同或相似的上游信号来激活LATS1/2?
2.在哺乳动物细胞中,Hippo通路成分的细胞定位在哪里?这可能是了解Hippo通路如何响应上游信号进而被调节的关键。很明显,磷酸化的YAP/TAZ富集在细胞质中,去磷酸化的YAP/TAZ富集在细胞核中。然而,不太清楚YAP/TAZ的磷酸化和去磷酸化发生在哪里。一个相关的问题是,YAP/TAZ在细胞核和细胞质之间转位的机制是什么?
3.LATS1/2如何受肌动蛋白重塑和/或细胞张力的调节?这是理解Hippo激酶级联调节的关键生化机制之一。越来越多的证据表明,肌动蛋白细胞骨架和细胞张力在LATS1/2的调控中起着关键作用,似乎在许多上游信号的下游起作用(如果不是大多数)。肌动蛋白细胞骨架和细胞张力是交织在一起的,那么问题来了:哪一个在调控Hippo核心成分中起着更直接的作用?
4.器官大小感应的机制是什么?尽管有许多信号被报道可在体外调节Hippo通路,但到目前为止,还没有一个信号通路被证明在体内器官大小的控制中起关键作用。找到这个神奇的信号将会解决发育生物学中的一个关键问题。
5.YAP在癌症中是如何变得失调的?虽然Hippo通路的基因突变很少见,但在很多种人类癌症中都观察到了YAP/TAZ的激活。这个表明,Hippo通路可能受到许多其他癌症驱动通路的广泛调节。
—END—
: , 。 视频 小程序 赞 ,轻点两下取消赞 在看 ,轻点两下取消在看
- 搜索
-
- 1000℃Nutrients:真实世界数据,纤维肌痛患者的饮食与运动自适应规律
- 1000℃D-二聚体升高诊治与管理专家共识(2026)
- 1000℃专家论坛|文良志:门静脉血栓的诊断和治疗
- 1000℃首例儿童NF2驱动型胸膜间皮瘤,多方法学检测锁定NF2双等位基因失活和14/22号染色体缺失,提示与成人胸膜间皮瘤不同
- 1000℃打破误区:干扰素追求CHB功能性治愈,HBsAg为何“不降反增”?
- 1000℃迷惑性极强的肝内病灶!影像表现疑点重重,最终病理竟查出两种不同肝脏恶性肿瘤
- 1000℃指南共识|原发性肝癌分子靶向药物相关蛋白尿中西医结合诊疗专家共识
- 1000℃Diabetologia:意大利北部社区 1~100 岁人群胰岛自身抗体与乳糜泻 TGA-IgA 的年龄分布及检测方法学验证
- 精J Child Psychol Psychiatry:12种罕见神经发育障碍儿童沟通能力谱系
- 精研究发现:爱吃辣的人,心血管病和癌症死亡风险都会显著降低
- 精Nursing in Critical Care:别再指责护士了!ICU 里被遗漏的护理,根源在系统而非个人
- 精Acta Obstet Gynecol Scand:罕见病女性的妊娠并发症与母婴结局,一项单中心434种罕见病的回顾性队列研究
- 精【爱儿小醉】儿科患者术前对流层臭氧暴露与围手术期呼吸系统不良事件之间的关系:一项单中心回顾性队列研究
- 精eBioMedicine:牙龈下微生物组与脑健康存在连续关联梯度,牙周炎或成认知衰退可干预靶点
- 精军事医学研究院《自然·通讯》:自适应IrPtCu纳米酶水凝胶实现耐药菌感染伤口序贯治疗
- 精能够逆转萎缩性胃炎的两个中成药,该怎么选择?
- 荐同时性多发性原发性肺癌,左右病灶分别为EGFR和ALK阳性,考虑淋巴结肿大仅局限左肺门及血浆EGFR阳性,采用奥希替尼联合化疗
- 荐40岁女性同时罹患卵巢支持细胞-间质细胞瘤和透明细胞乳头状肾肿瘤,WES等基因检测竟为阴性
- 荐椎管内麻醉使用止血药突发气道痉挛的抢救流程解析
- 荐女子肝区无任何不适,影像提示复杂囊性病变,层层鉴别后锁定罕见胆管源性囊性肿瘤
- 荐“绘”真报告 | 病理考虑为中枢神经细胞瘤,检出脑室外神经细胞瘤的特征性变异FGFR1-TACC1融合,辅助鉴别诊断
- 荐8岁女童出现男性化症状,竟是形似「性索-间质肿瘤」的卵巢「无性细胞瘤」所致,少见KRAS/CDK4共扩增或与侵袭性有关
- 荐17例病例分析揭示常见于中年人的色素性室管膜瘤临床特征与预后,分子检测可助力临床精准诊疗
- 荐Lancet Oncol:结直肠癌腹膜转移,围手术期化疗并非必选项
- 标签列表
-
- 星座 (702)
- 孩子 (526)
- 恋爱 (505)
- 婴儿车 (390)
- 宝宝 (328)
- 狮子座 (313)
- 金牛座 (313)
- 摩羯座 (302)
- 白羊座 (301)
- 天蝎座 (294)
- 巨蟹座 (289)
- 双子座 (289)
- 处女座 (285)
- 天秤座 (276)
- 双鱼座 (268)
- 婴儿 (265)
- 水瓶座 (260)
- 射手座 (239)
- 不完美妈妈 (173)
- 跳槽那些事儿 (168)
- baby (140)
- 女婴 (132)
- 生肖 (129)
- 女儿 (129)
- 民警 (127)
- 狮子 (105)
- NBA (101)
- 家长 (97)
- 怀孕 (95)
- 儿童 (93)
- 交警 (89)
- 孕妇 (77)
- 儿子 (75)
- Angelababy (74)
- 父母 (74)
- 幼儿园 (73)
- 医院 (69)
- 童车 (66)
- 女子 (60)
- 郑州 (58)